Иммунная система
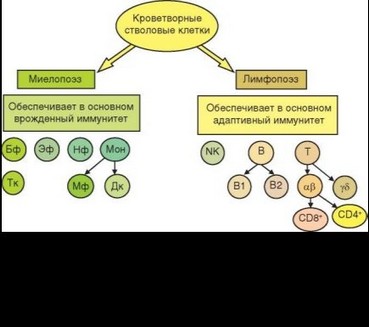
Иммунные процессы осуществляются клетками костномозгового происхождения, относящимися к двум кроветворным линиям — миелоидной и лимфоидной. Миелоидные клетки «отвечают» за реакции врожденного, лимфоидные — преимущественно за реакции адаптивного и только частично — врожденного иммунитета (рис. 1.2).
Врожденный иммунитет реализуется клетками (преимущественно фагоцитами), практически не нуждающимися в межклеточных контактах и коммуникациях. В связи с этим отсутствует необходимость их локализации в специализированных органах иммунной системы: амебоциты беспозвоночных и миелоидные клетки позвоночных широко распределены по организму; особенно богаты ими барьерные ткани. До формирования адаптивного иммунитета (например, у беспозвоночных) специальные органы иммунной системы отсутствовали.
Адаптивный иммунный ответ основан на постоянных межклеточных контактах и кооперации между клетками. Кроме того, в связи с клональной природой ответа возникает необходимость в особых механизмах концентрации (рекрутирования) клеток конкретных клонов в определенном месте. Обеспечение диалога между клетками и их вовлечение в иммунный ответ возможно лишь в условиях органной структуры. Поскольку адаптивный иммунный ответ обеспечивается лимфоидными клетками, органы иммунной системы являются прежде всего лимфоидными органами.
Иммунная система состоит из центрального и периферического отделов (рис. 1.3). Центральный отдел содержит органы (первичные, или цент-
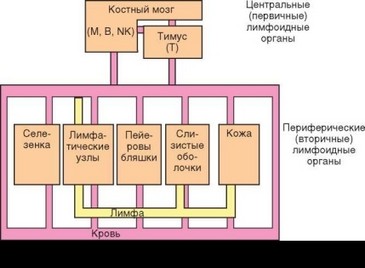
ральные, лимфоидные органы), в которых происходят дифференцировка и созревание лимфоцитов: костный мозг и тимус (вилочковая железа). Костный мозг — основной орган лимфо- и гемопоэза. Кроме того, он служит местом сосредоточения эффекторных клеток адаптивного иммунитета (например, плазмоцитов). Таким образом, только тимус является чисто центральным лимфоидным органом. Основная его функция — обеспечение развития Т-лимфоцитов (см. ниже). У птиц, некоторых рептилий, а также у жвачных млекопитающих имеется особый тип центральных лимфоидных органов или лимфоидных структур, в которых развиваются В-лимфоциты (у птиц и рептилий — сумка, или бурса Фабриция). Название центральных лимфоидных органов определило обозначение основных типов лимфоцитов: Т (тимусзави- симые) и В (бурсазависимые). Название третьего типа лимфоидных клеток — NK-клеток — происходит от выполняемой ими функции (естественные киллеры — natural killers).
Периферический отдел иммунной системы образован вторичными (периферическими) лимфоидными органами: неинкапсулироваными лимфоидными структурами, связанными со слизистыми оболочками, диффузно распределенными лимфоидными и миелоидными клетками и инкапсулированными (т.е. истинными, морфологически изолированными) лимфоидными органами. Лимфоидные органы взаимосвязаны путями рециркуляции лимфоцитов (лимфатическая и кровеносная системы). Выделяют 3 разновидности инкапсулированных лимфоидных органов — лимфатические узлы, селезенка и пейеровы бляшки. Все лимфоидные органы организованы сходным образом. В качестве примера можно рассмотреть структуру лимфатических узлов (см. рис. 3.68). В них выделяют зоны сосредоточения Т-лимфоцитов (паракортикальные зоны), В-лимфоцитов (фолликулы), а также сегменты, в которых В- и Т-клетки соседствуют друг с другом. В селезенке аналогично структурирована белая пульпа. Собственно, именно белая пульпа представляет вторичный лимфоидный орган, тогда как красная пульпа имеет иную структуру и ее функции только частично относятся к иммунитету. Аналог лимфатических узлов — пейеровы бляшки кишечника, содержащие структуры, связанных с транспортом антигенов через эпителиальный барьер; их главным компонентом являются эпителиальные М-клетки. Лимфоидные образования слизистых оболочек (миндалины, одиночные фолликулы, аппендикс) соответствуют отдельным структурным элементам лимфоидных органов, чаще всего лимфоидным фолликулам. Комплекс этих образований вместе с диффузно распределенными лимфоцитами, пейеровыми бляшками и региональными лимфатическими узлами формирует лимфоидную ткань, связанную со слизистыми оболочками (Mucosa-associated lymphoid tissue — MALT). Стромальные клетки лимфоидных органов и MALT способны привлекать клетки соответствующих типов и поддерживать их жизнеспособность, т.е. формируют для них нишу.
Завершив развитие в костном мозгу, миелоидные клетки поступают в кровь и некоторое время (обычно короткое) циркулируют в кровотоке. Из кровотока они мигрируют в ткани, в которых живут от нескольких суток до месяцев или лет. Кроме такого конститутивного пути миграции, существует экстренная миграция клеток (в основном миелоидных) из кровотока в места контакта с патогеном и очаг воспаления. Миелоидные клетки, участвующие в иммунных процессах, представлены моноцитами, нейтрофильными, эозинофильными и базофильными гранулоцитами. Некоторые разновидности миелоидных клеток практически не выявляются в кровотоке (хотя они тоже проходят стадию циркуляции), но присутствуют в тканях: тучные клетки и 2 типа тканевых клеток, образующихся из моноцитов, — макрофаги и дендритные клетки. Последние играют роль посредника между врожденным и адаптивным иммунитетом. Миелоидные клетки экспрессируют комплекс рецепторов, распознающих PAMP.
Как уже было упомянуто, выделяют 3 основных типа лимфоцитов — Т-, В- и естественные киллеры. NK-клетки относят к клеткам врожденного иммунитета. Естественные киллеры распознают молекулы (стрессорные молекулы), отличные от распознаваемых миелоидными клетками (PAMP). Как отмечалось, В- и Т-лимфоциты распознают антигены, однако это распознавание происходит по-разному. Иммуноглобулиновый рецептор В-клеток (BCR) дает им возможность распознавать нативный антиген как в свободной, так и в связанной с мембранами формах. Рецептор Т-клеток (TCR) распознает только фрагменты антигена, связанные с молекулами MHC. В процессе дифференцировки в Т- и В-лимфоцитах происходит перестройка генов, кодирующих рецепторы для антигенов. В результате каждая клетка экспрессирует рецептор, уникальный по специфичности. Рецепторы такой же специфичности имеют все потомки этой клетки (клон). В процессе селекции погибает большинство опасных аутоспецифических клонов как Т-, так и В-клеток. Популяции Т- и В-лимфоцитов участвуют в иммунных реакциях клонального типа, при которых в ответ вовлекаются только клетки клонов, экспрессирующих рецепторы нужной специфичности (в отличие от естественных киллеров, не отличающихся друг от друга по специфичности).
Популяции лимфоцитов гетерогенны не только по структуре анти- генраспознающего рецептора. К естественным (т.е. формирующимся в процессе нормальной дифференцировки, не связанной с действием чужеродных антигенов) относят 3 субпопуляции В-клеток. В1-клетки локализованы в серозных полостях и барьерных тканях, несут рецептор с низкой специфичностью к антигену, спонтанно вырабатывают низкоаффинные антитела преимущественно IgM-изотипа, в том числе к аутоантигенам. В-клетки маргинальной зоны — клетки, сходные с В1, но локализующиеся в маргинальной зоне селезенки. В2-клетки, которые мы привыкли называть обычными В-клетками, локализованы в селезенке и лимфатических узлах (в том числе в фолликулах), костном мозгу, лимфоидных тканях кишечника; эти клетки отвечают за образование высокоспецифичных и высокоаффинных антител разных изотипов.
Число естественных субпопуляций Т-лимфоцитов значительно больше. Прежде всего это — у5Т и арТ-клетки, отличающиеся типом TCR, а следовательно специфичностью распознавания и спектром функций. Среди арТ-клеток выделяют NKT-лимфоциты, совмещающие большинство функций NK клеток и некоторые функции Т-лимфоцитов. От обычных арТ-клеток они отличаются ограниченностью репертуара специфичностей TCR и преимущественным участием в распознавании липидных (а не пептидных) эпитопов. Среди «классических» арТ-клеток выделя-
ют субпопуляции CD4+ и CD8+ Т-клеток, отличающиеся двумя основными особенностями — распознаванием антигенных пептидов в составе разных молекул MHC (соответственно классов II и I) и функцией: после стимуляции антигеном CD4+ Т-клетки выступают в качестве хелперов, а CD8+ Т-клетки — в качестве цитотоксических Т-лимфоцитов. Если CD8+ Т-клетки в процессе иммунного ответа функционируют как единая субпопуляция, то среди CD4+ клеток выделяют несколько «адаптивных» субпопуляций, о которых будет сказано ниже. Однако существует одна естественная субпопуляция CD4+ Т-клеток, существенно отличающаяся от остальных Т-хелпе- ров — естественные регуляторные Т-клетки. Их функция состоит в контроле активности аутоспецифических клонов Т-лимфоцитов, не удаленных в процессе отрицательной селекции, мигрировавших в периферический отдел иммунной системы и создающих опасность аутоагрессии.
Лимфоциты, особенно Т-клетки, постоянно рециркулируют — выходят из лимфоидных органов в лимфу, мигрируют с ней в кровоток и возвращаются через посткапиллярные венулы обратно в орган. При этом благодаря экспрессии молекул адгезии и рецепторов для хемокинов (хемотаксических факторов, определяющих направление миграции клеток) рециркулирующие клетки при каждом «витке» рециркуляции с высокой избирательностью попадают в участки лимфоидных органов, специализированные для этого типа клеток. Некоторые миелоидные и лимфоидные клетки (в особенности ранее контактировавшие с антигеном) диффузно распределяются в барьерных и в меньшей степени — в других нелимфоидных тканях.
Клетки иммунной системы существенно различаются по сроку жизни. В соответствии с этим варьирует скорость их обновления. Численность клеток каждого типа строго контролируется гомеостатическими механизмами.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Иммунная система »
- Зарождение иммунологии
- Развитие иммунологии до середины ХХ века
- «Новая иммунология» 50-80-х годов ХХ века
- Современный этап развития иммунологии — молекулярная иммунология
- ЕСТЕСТВЕННАЯ ИСТОРИЯ ИММУНИТЕТА
- КРАТКОЕ ИЗЛОЖЕНИЕ ИММУНОЛОГИИ
- Молекулы-мишени иммунитета (образы патогенности, антигены) и распознающие их рецепторы
- Первая линия иммунной защиты
- Адаптивный иммунный ответ
- Эффекторные механизмы иммунного ответа. Взаимосвязь факторов врожденного и адаптивного иммунитета
- Иммунологическая память
- РЕЗЮМЕ