ЕСТЕСТВЕННАЯ ИСТОРИЯ ИММУНИТЕТА
Филогенез иммунитета неотделим от истории возникновения и развития многоклеточных организмов. Возникновение Metazoa (многоклеточные) означает формирование автономных организмов, имеющих внутреннюю среду, заполненную принадлежащими данному организму клетками и ограниченную барьером, отделяющим ее от окружения. Окружение a priori враждебно организму, поскольку служит источником агрессии, конкуренции и т.д. Агрессия может состоять в проникновении других организмов (прежде всего одноклеточных) во внутреннюю среду многоклеточного организма с последующей конкуренцией за территорию и ресурсы, а также возможным активным повреждением клеток или их отравлением токсинами и метаболитами. Таким образом, сам факт возникновения обособленного сообщества клеток, имеющего хотя бы элементарные интегрирующие системы и воспроизводящегося как единое целое, послужило достаточным основанием для возникновения «службы» поддержания клеточного и молекулярного постоянства внутренней среды. Такая «служба» и стала прообразом иммунной системы.
Из сказанного выше следует, что первое условие формирования иммунитета — наличие «охраняемой» замкнутой территории с ее обязательным отграничением от внешней среды. Второе условие — появление факторов, специализированных для обеспечения постоянства охраняемой внутренней среды путем ее освобождения от поступивших извне агентов (т.е. для обеспечения иммунитета в его прямом первоначальном смысле — освобождение). Со времен И.И. Мечникова общепризнано, что таким фактором стали специализированные клетки мезенхимального происхождения — подвижные амебоциты, предки фагоцитов млекопитающих. Они обладают выраженной способностью к фагоцитозу — механизму, обеспечивающему элиминацию потенциально агрессивных клеток, проникших во внутреннюю среду организма.
Важное условие эффективной работы этого гомеостатического механизма — способность защитных клеток отличать потенциально агрессивные чужие клетки от собственных. Принцип, на который опирается такое распознавание, стал основой иммунитета во всех его проявлениях. Таким образом, иммунная система, не имея возможности «дожидаться» проявления агрессивности проникших извне клеток, рассматривает в качестве потенциально опасных любые чужеродные клетки и молекулы. По-видимому, такое «решение» эволюции наиболее универсально и оправдано: действительно чужеродные объекты практически всегда вредны, даже если они не проявляют активной агрессии.
Возникновение рецепторов, позволяющих «опознать» чужое, стало третьим основополагающим событием на пути формирования иммунитета (после возникновения внутренней среды многоклеточных и специализированных клеток-фагоцитов). Действительно, наличие патогенраспоз- нающих рецепторов, как их теперь называют, — чрезвычайно древнее «изобретение» эволюции, общее для животных и растений. Отметим сразу, что иммунитет растений и животных эволюционировал в последующем разными путями, однако общий принцип распознавания чужеродных объектов сохранился.
В процессе эволюции вида закреплялись гены, кодирующие молекулы, предназначенные для распознавания не просто «чужого», но заведомо опасного для данного организма. Эти рецепторы — мембранные или растворимые молекулы, обладающие пространственным сродством (и потому способные их распознавать) к наиболее общим и связанным с патогенностью молекулярным маркерам чужеродных агентов: компонентам клеточной стенки бактерий, эндотоксинам, нуклеиновым кислотам и т.д. Каждый рецептор распознает не индивидуальную молекулу, а целую группу подобных молекул, служащих образами (паттернами) патогенности. Молекулы- рецепторы представлены не только на поверхности клеток-эффекторов иммунитета, но и в гранулах, в которые попадают чужеродные агенты при фагоцитозе. Патогенраспознающие молекулы присутствуют также в жидких средах организма и способны инактивировать токсины и убивать чужеродные клетки. Относительно небольшое число генов, кодирующих такие рецепторы, обеспечивает распознавание практически всех патогенов, не составляя чрезмерной «обузы» для многоклеточного организма.
В результате распознавания образов патогенности происходит активация клеток — иммуноцитов, что позволяет им убивать и затем элиминировать патогены. Это происходит с помощью цитолиза — внутриклеточного (наиболее совершенного, связанного с фагоцитозом), внеклеточного (вызываемого секретируемыми факторами) и контактного. Патогены могут быть убиты или подготовлены к фагоцитозу растворимыми бактерицидными факторами и рецепторными молекулами. Во всех случаях окончательное расщепление убитых патогенов происходит в процессе фагоцитоза.
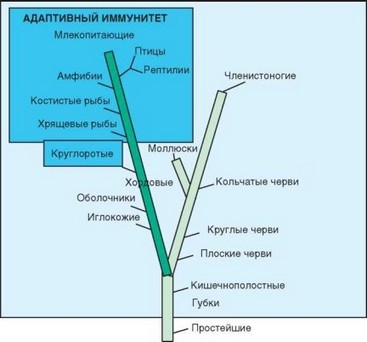
Рис. 1.1. Филогенез врожденного и адаптивного иммунитета. На упрощенном филогенетическом древе (обозначены только те таксоны, у которых исследовали иммунитет) отмечены зоны действия врожденного и адаптивного иммунитета. Круглоротые выделены в особую группу как животные, у которых адаптивный иммунитет развивался не по «классическому» пути
Так, схематично можно представить систему иммунитета, которую принято называть врожденной. Эта форма иммунитета характерна для всех многоклеточных животных (в несколько иной форме — и для растений). Ее возраст — 1,5 млрд лет. Система врожденного иммунитета весьма эффективно защищала первичноротых многоклеточных животных, а также низших вторичноротых, часто имевших крупные размеры (рис. 1.1). Проявления врожденного иммунитета на разных стадиях эволюции и в разных таксонах чрезвычайно разнообразны. Однако общие принципы его функционирования одинаковы на всех стадиях развития многоклеточных. Главные составляющие врожденного иммунитета:
- распознавание чужеродных агентов во внутренней среде организма с помощью рецепторов, специализированных на узнавании «образов» патогенности;
- элиминация опознанных чужеродных агентов из организма путем фагоцитоза и расщепления.
У хордовых произошло скачкообразное формирование другой разновидности иммунитета: примерно 500 млн лет назад возник адаптивный (т.е. приспособительный) или приобретенный иммунитет. Ветвь адаптивного иммунитета, получившая интенсивное развитие, зародилась у хрящевых рыб. Особый вариант адаптивного иммунитета, основанный на использовании других распознающих и эффекторных молекул, обнаружен у более примитивных хордовых — круглоротых. Адаптивный иммунитет тесно связан с врожденным и во многом основывается на его проявлениях. Однако эти типы иммунитета сильно различаются (табл. 1.2).
Таблица 1.2. Основные свойства врожденного и адаптивного иммунитета
|
Характеристика |
Врожденный иммунитет |
Адаптивный иммунитет |
|
Условия формирования |
Формируется в онтогенезе вне зависимости от «запроса» |
Формируется в ответ на «запрос» (поступление чужеродных агентов) |
|
Объект распознавания |
Группы чужеродных молекул, связанных с патогенностью |
Индивидуальные молекулы (антигены) |
|
Эффекторные клетки |
Миелоидные, частично лимфоидные клетки |
Лимфоидные клетки |
|
Тип реагирования популяции клеток |
Популяция клеток реагирует как целое (не клонально) |
Реакция на антиген клональная |
|
Распознаваемые молекулы |
Образы патогенности; стрес- сорные молекулы |
Антигены |
|
Распознающие рецепторы |
Патогенраспознающие рецепторы |
Антигенраспознающие рецепторы |
|
Угроза аутоагрессии |
Минимальная |
Реальная |
|
Наличие памяти |
Отсутствует |
Формируется иммунологическая память |
Существенное отличие адаптивного иммунитета от врожденного — способ распознавания чужого (табл. 1.3). В адаптивном иммунитете оно осуществляется при помощи молекул особого типа (иммуноглобулинов или других белков суперсемейства иммуноглобулинов), при этом распознаются не паттерны, а индивидуальные молекулы или небольшие группы сходных молекул, называемые антигенами. Существует порядка 106 различных антигенов. Такое число рецепторов не только не может быть представлено на одной клетке, но и не может быть закодировано в геноме позвоночных, содержащем только десятки тысяч генов. Именно поэтому в процессе эволюции адаптивного иммунитета сформировался сложный механизм генерации разнообразия антигенспецифических рецепторов: при развитии специализированных клеток (лимфоцитов), происходит перестройка их генов, кодирующих антигенраспознающие рецепторы, что приводит к образованию в каждой клетке рецептора с уникальной специфичностью. При активации каждая клетка может дать начало клону, все клетки которого будут иметь рецепторы той же специфичности. Таким образом, каждый конкретный антиген распознают не все лимфоциты, а только отдельные их клоны, имеющие специфические антигенраспознающие рецепторы.
Таблица 1.3. Основные типы иммунологического распознавания
|
Характеристика |
Групповое (паттерновое) |
Индивидуальное (антигенное) |
|
Объект распознавания |
Консервативные молекулярные структуры — образы патогенности |
Антигенные эпитопы (в составе свободных молекул или встроенные в молекулы MHC) |
|
Дискриминация «свое—чужое» |
Совершенная, сложилась в филогенезе |
Несовершенная, формируется в онтогенезе |
|
Потребность в костимуляции |
Нет |
Есть |
|
Время реализации эффекта |
Немедленно |
Требует времени (адаптивный иммунный ответ) |
|
Связь с различными формами иммунитета |
Связано с врожденным иммунитетом |
Связано с адаптивным иммунитетом |
|
Формирование генов рецепторов |
Детерминированы генетически |
Формируются в процессе дифференцировки клеток |
|
Клетки, несущие рецепторы |
Любые ядерные клетки (преимущественно миелоидные) |
Только В- и Т-лимфоциты |
|
Распределение на клетках |
Все клетки в популяции экспрессируют одинаковые рецепторы |
Клональное |
|
Рецепторы |
TLR, NLR, CLR, RIG, DAI, Seavenger-рецепторы, растворимые рецепторы |
BCR (на В-клетках), TCR-yS, (на у8Т-клетках), TCR- ар (на арТ-клетках) |
Если паттернраспознающие рецепторы врожденного иммунитета образовались в процессе эволюции как молекулы, распознающие чужеродные, но не собственные молекулы организма, то специфичность антигенрас- познающих рецепторов системы адаптивного иммунитета формируется случайно. Это потребовало развития дополнительных механизмов селекции для устранения «ненужных» и «опасных» (направленных против «своего») клонов лимфоцитов. Такие механизмы достаточно эффективны, однако все же не полностью устраняют риск развития аутоиммунных процессов — иммунных реакций, направленных против собственных антигенов, вызывающих повреждение организма хозяина.
Оба типа иммунитета образуют целостную систему, при этом врожденный иммунитет служит фундаментом для развития адаптивного. Так, лимфоциты распознают антиген в процессе презентации, осуществляемой преимущественно клетками врожденного иммунитета. Удаление из организма антигена и несущих его клеток происходит с помощью реакций, в основе которых лежат механизмы врожденного иммунитета, получившие специфический компонент, т.е. направленные на конкретный антиген и действующие с повышенной эффективностью.
Клональный характер адаптивного иммунного ответа создал возможность возникновения иммунологической памяти. При врожденном иммунитете память не развивается и каждый раз реакции на внедрение чужерод
ных молекул развиваются как впервые. В процессе адаптивного иммунитета формируются клоны клеток, сохраняющих «опыт» предыдущего иммунного ответа, что позволяет им реагировать на повторную встречу с антигеном значительно быстрее, чем при первичном контакте, и формировать при этом более сильный ответ. Наличие клеток памяти делает организм устойчивым к довольно широкому кругу патогенов. Вероятно, именно возможность формирования иммунологической памяти послужила преимуществом, позволившим закрепиться в процессе эволюции такому «дорогостоящему» для организма, громоздкому, во многом ненадежному и даже опасному механизму, как адаптивный иммунный ответ.
Таким образом, адаптивный иммунитет базируется на трех главных процессах:
- распознавании антигенов (как правило, чужеродных для организма) независимо от их связи с патогенностью, с помощью клонально распределенных рецепторов;
- элиминации распознанных чужеродных агентов;
- формировании иммунологической памяти о контакте с антигеном, позволяющей быстрее и эффективнее удалять его при повторном распознавании.
Адаптивный иммунитет имеет еще одно преимущество, отсутствующее у врожденного иммунитета — способность защищать организм от агрессии изнутри (т.е. от злокачественных новообразований). Риск развития злокачественных опухолей вследствие мутаций или вирусной трансформации клеток существенно возрос при увеличении в эволюции размеров организма, произошедшем примерно тогда же, когда возник адаптивный иммунитет. Помимо этого нельзя исключить, что адаптивный иммунитет возник как частное проявление изменений более высокого порядка, с которыми связаны существенные эволюционные преимущества, раскрыть которые предстоит в будущем.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «ЕСТЕСТВЕННАЯ ИСТОРИЯ ИММУНИТЕТА »
- Зарождение иммунологии
- Развитие иммунологии до середины ХХ века
- «Новая иммунология» 50-80-х годов ХХ века
- Современный этап развития иммунологии — молекулярная иммунология
- КРАТКОЕ ИЗЛОЖЕНИЕ ИММУНОЛОГИИ
- Молекулы-мишени иммунитета (образы патогенности, антигены) и распознающие их рецепторы
- Иммунная система
- Первая линия иммунной защиты
- Адаптивный иммунный ответ
- Эффекторные механизмы иммунного ответа. Взаимосвязь факторов врожденного и адаптивного иммунитета
- Иммунологическая память
- РЕЗЮМЕ