Развитие арТ-лимфоцитов
При значительном сходстве путей развития В- и Т-лимфоцитов диффе- ренцировка Т-клеток более продолжительна (табл. 3.13). Это объясняется главным образом особенностями распознающего аппарата Т-клеток, требующего сложной «настройки». Т-лимфоциты — единственные клетки крови, развитие которых не может осуществляться в микроокружении костного мозга — основные этапы дифференцировки они проходят в специальном органе — вилочковой железе, или тимусе. Тимус содержит эпителиальные клетки, отсутствующие в костном мозгу. Они и являются тем уникальным и ключевым фактором микроокружения, который способен обеспечить развитие Т-лимфоцитов. Структура тимуса и его роль в развитии Т-клеток будут рассмотрены далее (см. раздел 3.4.1.2).
Таблица 3.13. Стадии развития и субпопуляции тимоцитов человека
|
Стадия развития |
Фенотип |
Содер- жание,% |
Локализация |
Функция, процессы |
|
Дважды отрицательные (DN) |
Стадия DN1: CD4-CD8- CD3-; CD44+ CD25- |
4-5 |
Кортикомедуллярная и перимедуллярная зона |
Полипотентные предшественники |
|
Стадия DN2 (про-Т-клетки): CD4-CD8- CD3-; CD44+ CD25+ |
Субкапсулярная зона |
Подготовка перестройки генов |
||
|
Стадия DN3 (пре-Т-клетки): CD4- CD8- CD3- CD44- CD25+ |
То же |
Перестройка Vp-гена |
||
|
Незрелые моноположительные (iSP) |
CD4+ CD8- CD3- |
О I/i о Vi |
Наружные слои коры |
Нет данных |
|
Дважды положительные (DP) TCRap+ |
CD4+ CD8+ CD3-TCRaplo CD69-CD5lo |
70-75 |
Кора |
Перестройка Va-гена. Незрелые Т-клетки; положительная селекция |
|
CD4+ CD8+ CD3-TCRaphi CD69- CD5+ |
Глубокие слои коры |
Отрицательная селекция. Дифференцировка CD4/CD8 |
||
|
Моно-положи- тельные (SP) TCRap+ |
CD4+ CD8- CD3- TCRaphi CD25- |
6-7 |
Мозговой и кортикомедуллярный слои |
Предшественники Т-хелперов |
|
CD4- CD8+ CD3- TCRaphi |
4-5 |
То же |
Предшественники цитотоксических Т-лимфоцитов |
|
|
CD4+ CD8- CD3- TCRaphi CD25hi |
2-3 |
— lt;lt; — |
Естественные регуляторные Т-клетки |
|
|
Дважды отрицательные (DN) TCRyS+ |
CD4- CD8- CD3- TCRyS+ |
1 |
Наружные слои коры |
Предшественники Т-клеток барьерных тканей |
Миграция в тимус клеток-предшественников
Из кроветворных органов в тимус мигрируют ранние лимфоидные предшественники ELP (от Early lymphoid progenitors) — клетки с широким потенциалом дифференцировки, включая миелоидный путь. Фенотипически эти клетки можно охарактеризовать как CD34+ SCA-1+ CD117 (c-Kit)+ Flt-3+ CCR9+ CD410. Эта стадия развития предшествует стадии CLP. ELP отличаются от CLP отсутствием a-цепи рецептора для IL-7 и слабой экспрессией CD4. У человека фенотип этих клеток — CD34+ GD38l° CD45RA+ CD7+ (главное отличие от предшественников В-клеток состоит в экспрессии CD7 вместо CD10).
ELP попадают в тимус через венулы кортикомедуллярного сочленения тимуса (см. раздел 3.4.1.2), откуда они мигрируют в наружные слои коры. Затем начинается их возвратное движение в сторону кортикомедуллярного сочленения. В ходе этих перемещений происходит созревание Т-клеток. Уже на 1-м этапе (движение в сторону коры) клетки последовательно утрачивают способность дифференцироваться в В-лимфоциты, миелоидные клетки, естественные киллеры и дендритные клетки. Вероятно, на этом этапе внутри тимуса от клеток-предшественнков Т-линии ответвляются предшественники перечисленных «не-Т-клеток» (например, дендритных для обеспечения потребности в этих клетках внутри тимуса). На этом отрезке миграции проходят первые этапы развития самих Т-лимфоцитов.
Для маркирования стадий развития Т-лимфоцитов чаще используют экспрессию на их поверхности корецепторов CD4 и CD8, а также рецепторного комплекса CD3—TCR (см. рис. 3.44). Наиболее ранние представители Т-клеточного ряда имеют фенотип CD4-CD8-, в связи с чем их называют дважды отрицательными или дважды негативными (DN — от Doublenegative) клетками. Поскольку они лишены также CD3, эти клетки иногда называют трижды негативными клетками. Более зрелая популяция ав-тимоцитов экспрессирует оба корецептора и обозначается как дважды положительные клетки — CD4+CD8+ (дважды позитивные, DP — от Doublepositive). Они уже экспрессируют на своей поверхности (как правило, слабо) CD3. Наконец, зрелые тимоциты, как и периферические Т-клетки, экспрессируют один из корецепторов и поэтому их называют моноположительными клетками (SP — от Single-positive). Как уже упоминалось выше, фенотип CD4+CD8- характерен для Т-хелперов и регуляторных Т-клеток, а CD4-CD8+ — для цитотоксических Т-лимфоцитов.
Развитие Т-клеток сходно, но не идентично развитию В-лимфоци- тов. Как и в случае В-клеток, основные события при Т-лимфопоэзе — перестройка рецепторных V-генов и формирование TCR с последующей селекцией клонов Т-лимфоцитов, обеспечивающей формирование анти- генраспознающего репертуара. Перестройка V-генов TCR и другие события, приводящие к экспрессии рецептора, происходят на стадии дважды отрицательных тимоцитов в поверхностных слоях коры тимуса (субкапсулярной зоне). Для удобства рассмотрения этого процесса в стадии дважды отрицательных DN (тимоцитов) выделяют подстадии по экспрессии двух мембранных молекул — CD44 (рецептор, распознающий гиалуронат и ответственный за миграцию клеток-предшественников в тимус) и CD25 (а-цепь рецептора для IL-2). DNl-клетки, мигрирующие в тимус, имеют фенотип CD44+ CD25-, DN2 — CD44+ CD25+, DN3 — CD44- CD25+.
Развитие дважды отрицательных тимоцитов и перестройка генов T-клеточного рецептора (см. табл. 3.13)
Самые ранние CD44+ CD25- тимоциты локализуются в перимедуллярной зоне коры тимуса. Они представляют собой полипотентные кроветворные предшественники, их V-гены имеют зародышевую, т.е. неперестроенную конфигурацию. Только в некоторых клетках на этой стадии проходит начальный, малоспецифичный этап перестройки генов — сближение сегментов D и J в генах в- и 5-цепей (см. рис. 3.43). На стадии DN1 клетки
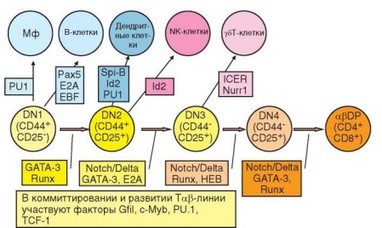
Рис. 3.45. Полипотентность ранних тимических предшественников и роль диффе- ренцировочных факторов в выборе пути их развития. Клетки-предшественники, поступающие в тимус, обладают широким дифференцировочным потенциалом, который они в процессе развития постепенно утрачивают. В конце стадии дважды отрицательных клеток тимоциты сохраняют способность дифференцироваться только в арТ-клетки
интенсивно пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению их числа. Это наиболее продолжительный этап развития Т-клеток (10 сут — 50% времени развития Т-клеток в тимусе). Основной ростовой фактор на этом этапе — SСF (С-Kit лиганд). В это время рецептор для IL-7 экспрессирован слабо или вообще не экспрессирован. Факторами дифференцировки на этом этапе развития служат Wnt и Hedgehog, а также Notch, коммиттирующий клетки-предшественники к дифференцировке в Т-лимфоциты. Дальнейшие стадии дифференцировки Т-клеток представлены на рис. 3.45.
На стадии DN2 тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого эпителиальными клетками наружной коры. Длительность стадии — 2 сут. Клетки продолжают пролиферировать уже преимущественно под действием IL-7 и в меньшей степени — SCF. На этой стадии сигналы от Notch коммиттируют Т-клетки к дифференци- ровке в различные линии (у5 или ав) и экспрессируется ген, кодирующий pTCRa, — знак выбора ав-направления развития. Это коммиттирование становится необратимым только после реаранжировки генов TCR на стадии DN3. Тогда же экспрессируются гены рекомбинационного комплекса.
При переходе на стадию DN3 запускается основное событие дифференцировки Т-лимфоцитов — перестройка V-генов TCR. Включение этого процесса является следствием экспрессии в тимоцитах группы дифферен- цировочных факторов под влиянием эпителиального микроокружения. Для раннего развития Т-клеток необходима экспрессия генов Notch, Runx-1,
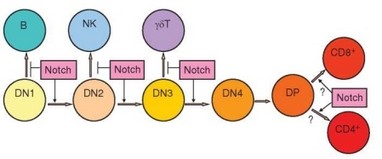
Рис. 3.46. Роль Notch-рецепторов в развитии Т-клеток в тимусе. Показано стимулирующее влияние Notch на разные стадии развития арТ-клеток в тимусе и ингибирующее — на альтернативные пути дифференцировки
GATA-3, Ikaros, Gfi1, c-Myb, PU.1, E2A/HEB, TCF-1. Особенно важную роль играют факторы семейства Notch (особенно Notch-1 и Notch-3), действующие почти на всех этапах раннего развития тимоцитов и блокирующие другие пути дифференцировки клеток-предшественников (в направлении В-, NK-, дендритных клеток) (рис. 3.46). Механизм действия этого основного дифференцировочного фактора Т-клеток представлен на рис. 3.47. Для запуска перестройки V-генов TCR наиболее важна передача сигнала, осуществляемая Notch-1 на стадиях DN2 и DN3. Рецептор для этого фактора экспрессируется тимоцитами на указанных стадиях развития. Лиганды Notch-рецептора — молекулы Delta и Jagged — находятся на поверхности
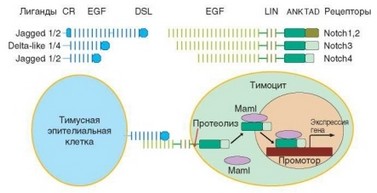
Рис. 3.47. Notch-рецепторы, их лиганды и Notch-опосредованная передача сигнала. Схема иллюстрирует строение Notch-рецепторов и Notch-лигандов (основные компоненты их внеклеточной части — EGF-подобные повторы). Показано гомотипическое взаимодействие лиганда и рецептора, освобождение внутриклеточной части рецептора вследствие протеолиза и ее функционирование в качестве ядерного транскрипицонного фактора
эпителиальных клеток тимуса. Определенную роль в индукции перестройки V-генов играют цитокины, в частности IL-7, который особенно важен для запуска перестройки V-гена у-цепи. Еще более существенна роль IL-7 в качестве фактора выживания, индуцирующего экспрессию антиапоптоти- ческого фактора Bcl-2. На этом этапе развития после запуска перестройки V-генов TCR развитие макрофагов и В-лимфоцитов становится невозможным.
Реаранжировка происходит в основном при переходе от стадии DN2 (CD4+ CD25+) к стадии DN3 (CD44- CD25+), которые называют также стадиями про-Т и пре-Т, соответственно (см. рис. 3.43). На этих стадиях экспрессируются гены RAG1 и RAG2, ген фермента TdT и гены других компонентов рекомбинационного комплекса. Перестройка V-генов TCR происходит, как принято считать, в последовательности 5, у, р. Однако это правило соблюдается не очень строго: все 3 гена начинают перестраиваться практически одновременно. Как и в случае V-генов иммуноглобулинов, сначала перестраиваются гены только на одной хромосоме. В генах р- и 5-цепей сближаются сегменты D и J, а затем происходит полномасштабная перестройка с вовлечением основного сегмента V-гена и формированим непрерывных последовательностей VDJ. Реаранжировка генов у-цепи проходит так же, но с одним отличием: поскольку в гене у-цепи отсутствует D-сегмент, в результате перестройки формируется последовательность VJ. Как и при реаранжировке V-генов иммуноглобулинов, при неудачной перестройке V-гена на одной хромосоме процесс повторяется на другой хромосоме. Успешная перестройка служит сигналом к прекращению этого этапа реаранжировки. Это обусловливает экспрессию клеткой только одного варианта продуктов перестроенных V-генов. Реаранжировка V-генов TCR делает невозможной дифференцировку про-Т-клеток в направлении NK- и дендритных клеток.
При удачной перестройке V-гена p-цепи экспрессируется пре-Г-рецеп- тор, аналогичный рассмотренному выше пре-В-рецептору (см. рис. 3.38). Он представляет собой димер, состоящий из полноценной p-цепи и инвариантной (т.е. лишенной вариабельного участка) цепи, называемой пре-TCRa. Экспрессия этой молекулы на мембране сопровождается ее спонтанной олигомеризацией, что само по себе (т.е. без связывания какого-либо лиганда) сигнализирует о благополучной перестройке. Экспрессия пре-Г-рецеп- тора и передача сигнала с этой молекулы служит 1-й контрольной точкой перестройки TCR. Ответ на этот сигнал — усиление экспрессии фактора, блокирующего развитие апоптоза (Вс1-2), и запуск пролиферации клеток (индуцируется преимущественно IL-7). Если после двух попыток перестройки проторецептор не экспрессируется, клетка подвергается апоптозу. Как и при реаранжировке V-генов иммуноглобулинов, успешная перестройка V-гена p-цепи происходит в 55% клеток. Отбор тимоцитов, успешно перестроивших V-ген p-цепи, путем подавления их апоптоза и индукции пролиферации, называют р-селекцией. В период р-селекции временно прекращается экспрессия генов RAG1 и RAG2 и приостанавливается процесс перестройки генов.
Сложным и не до конца понятным остается вопрос о механизмах выбора между у5 - и ap-путями дифференцировки Т-клеток (рис. 3.48). На стадии DN3, когда тимоцит экспрессирует мембранный пре-Г-рецептор, он уже
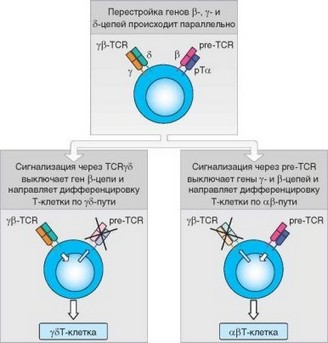
Рис. 3.48. Выбор пути ар/у5-дифференцировки Т-клеток
может нести на своей поверхности полноценный у5 -TCR. Известно, что каждая Т-клетка может экспрессировать TCR одного типа (ар или у8). Таким образом, на этапе DN3 клетка должна сделать выбор: продолжать перестройку V-гена а-цепи для формирования TCR ар-типа или довольствоваться уже сформированным y5TCR. В настоящее время наиболее широко принята концепция «силы сигнала», согласно которой, если y5TCR передают сигналы достаточной силы (заведомо превосходящей силу сигнала от пре-Т-рецепторов), Т-клетки прекращают перестройку V-генов и развиваются как у5Т-лимфоциты. В противном случае экспрессия генов у- и 5-цепей блокируется и клетки продолжают перестройку V-генов. Перестройка V-гена а-цепи делает физически невозможным развитие тимоцитов в направлении у5-клеток, т.к. гены а- и 5-цепи образуют единый локус (см. 3.1.4), и перестройка гена а-цепи приводит к удалению последовательностей, кодирующих 5-цепь.
По-видимому, микроокружение эмбрионального и взрослого тимуса существенно различается по способности поддерживать формирование субпопуляций Т-клеток у5- и ар-линий. В тимусе эмбрионов развиваются преимущественно у5Т-клетки, затем эмигрирующие в периферический отдел иммунной системы (см. раздел 3.4.1.2). В тимусе взрослых животных на долю у5Т-клеток приходится около 1% тимоцитов, что отражает безусловное преобладание ар-тимопоэза. Кроме того, у5-тимоциты по мере завершения перестройки генов TCR быстро эмигрируют из тимуса, а арТ-клетки продолжают развитие, подвергаясь селекции. Образование арТ-клеток во взрослом организме происходит практически только в тимусе и арТ-лим- фопоэз является основной функцией тимуса взрослых животных.
К концу фазы DN3 тимоциты приобретают фенотип CD44-CD25+. Длительность фазы DN3 до р-селекции составляет 2 сут, а период р-селек- ции — еще 1 сут. На заключительном этапе фазы DN3 на клетках экспрессируется корецептор CD4 (стадия ISP — от Immature single-positive, т.е. незрелых моноположительных тимоцитов).
Ранее полагали, что существует особая стадия развития DN-тимоцитов — DN4, однако оказалось, что тимоциты приобретают фенотип CD44-CD25- одновременно с экспрессией корецепторов CD4 и CD8 (точнее, экспрессией молекулы CD8 в дополнение к уже присутствующей на мембране молекулы CD4). Продолжительность стадии дважды положительных тимоцитов невелика (1,5—2 сут), однако в это время происходит несколько событий, принципиально важных для развития Т-лимфоцитов. Одно из этих событий — реализация завершающего этапа формирования арTCR — перестройка V-гена а-цепи. Поскольку в а-гене отсутствуют D-сегменты, реаранжировка приводит к образованию VJ-продукта. Хотя вероятность возникновения ошибки при этом почти такая же, как при перестройке V-гена р-цепи, результативность реаранжировки а-гена выше. При неудачной перестройке генов, расположенных на обеих хромосомах, процесс начинается заново, причем в него вовлекаются другие V- и J-сегменты. Такой «перебор» сегментов продолжается до успешной перестройки, что может занять 3—4 сут. Все это время не прекращаются экспрессии генов, кодирующих RAG-1, RAG-2 и TdT. Именно поэтому вероятность неудачи на этом этапе реаранжировки даже ниже, чем на этапе перестройки V-генов L-цепи иммуноглобулинов в В-клетках.
После перестройки V-генов обеих цепей происходит сборка их продуктов и экспрессия «зрелого» TCR на поверхности клетки. Важно, что специфичность TCR у каждого индивидуального тимоцита различна, поскольку процесс перестройки осуществляется в них независимо от других клеток и вероятность совпадения нуклеотидной последовательности в перестроенных генах разных клеток практически равна нулю. Зрелый TCR экспрессируется в комплексе с CD3 и Z-цепями. Поскольку CD3 и Z-димер отвечают за передачу сигнала от рецептора в ядро, клетки уже на этом этапе имеют функционально полноценный рецептор. Вероятно, появление на поверхности тимоцита зрелого рецептора служит сигналом, необходимым для прекращения экспрессии Т-клеткой генов RAG1 и RAG2 и для поддержания ее жизнеспособности. В этом состоит суть второй «контрольной точки» при формировании TCR. Плотность экспрессии TCR на поверхности тимоци- тов сначала невелика. Выживаемость клеток в этот период обеспечивается взаимодействием молекулы семейства TNF — CD70 тимоцита и молекулы семейства TNFR — CD27 эпителиальной клетки.
Источник: Ярилин.А.А , «Иммунология » 2010