Феномен внутрисосудистой агрегации форменных элементов крови
Этот феномен обычно называют агрегацией эритроцитов, хотя, без сомнения, агрегаты содержат также и тромбоциты, и лейкоциты. Однако, учитывая, что количество эритроцитов в единице объема крови и агрегатах на один-два порядка больше количества других форменных элементов, термин «агрегация эритроцитов» в целом соответствует действительности.
Механизмы возникновения и развития агрегации эритроцитов весьма сложны и многообразны. Они до сих пор уточняются. Вместе с тем среди этих механизмов уже теперь можно выделить те, которые имеют ведущее значение. Заслуга детального изучения феномена внутрисосудистой агрегации эритроцитов принадлежит М. Knisely (1947, 1965). Обширный фактический материал собственных исследований и анализ работ других авторов позволили ему утверждать, что в норме ни у людей, ни у животных этот феномен практически не обнаруживается (подразумевается прижизненная агрегация эритроцитов).
Агрегаты эритроцитов при их образовании в патологических условиях закупоривают мелкие сосуды, ухудшают нутритивный кровоток и, таким образом, неблагоприятно влияют на транскапиллярный обмен [Knisely М. et al., 1947]. Крайнюю степень агрегации эритроцитов принято обозначать термином «сладжинг» (sludging). Необходимо различать агрегацию эритроцитов и их агглютинацию. Агрегация — процесс обратимый, тогда как агглютинация всегда необратима и обусловлена обычно иммунными факторами.
Результаты исследований обсуждаемого феномена с использованием метода реоскопии послужили основанием для деления эритроцитарных агрегатов на патологические и физиологические [Goldstone Y. et al., 1970]. Было установлено, что патологические агрегаты резистентны к сдвигу и не распадаются, как физиологические, а, напротив, уплотняются. Их форма и ригидность также значительно отличаются от этих параметров физиологических агрегатов. Патологические агрегаты обычно очень быстро оседают.
Выделение понятия «физиологические агрегаты» не противоречит положению об отсутствии агрегации эритроцитов в норме, а лишь дает основание говорить о едином динамическом процессе агрегация—дезагрегация, который постоянно протекает в крови. В норме дезагрегация доминирует над агрегацией.
Установлено, что интенсивность процесса дезагрегации зависит от скорости деформации. При увеличении градиента скорости число агрегатов в крови, сначала крупных, а затем и мелких (не более 30 клеток в одном агрегате), постепенно уменьшается, а при критической скорости деформации наступает полная дезагрегация [Левтов В.А. и др., 1982; Schmid-Schonbein Н. et al., 1977]. Дезагрегация сопровождается увеличением текучести крови. Полное разрушение агрегатов наступает обычно при градиентах скорости 50—80 с-1. Это свидетельствует о преобладании при этих скоростях деформации гидродинамических сил потока, стремящихся разобщить эритроциты, над силами межэритроцитарного взаимодействия. Эритроциты при этом переориентируются в потоке, стремясь обеспечить минимум диссипации энергии при наибольшей «устойчивости» [Шадрина Н.Х., 1976; Chien S., 1970].
Агрегация эритроцитов — один из важных факторов, обусловливающих нелинейность кривой течения крови. При низких скоростях деформации вклад агрегации в абсолютные значения эффективной вязкости максимален.
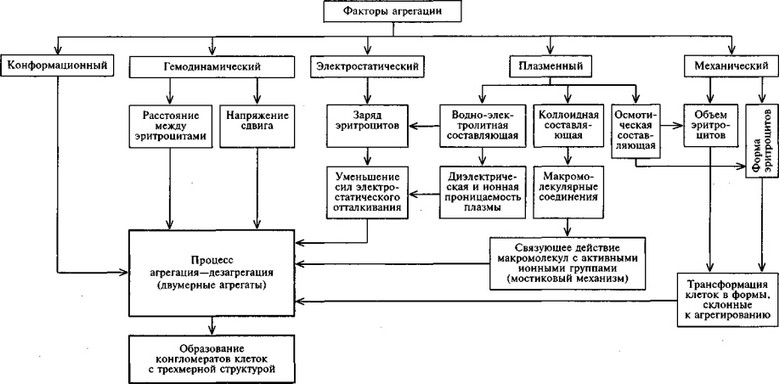
Результирующая направления процесса агрегация—дезагрегация в организме определяется взаимодействием по меньшей мере пяти факторов: гемодинамического, плазменного, электростатического, механического и конформационного.
Влияние гемодинамического фактора определяется зависимостью направления процесса агрегация—дезагрегация от напряжения сдвига и расстояния между отдельными клетками в потоке, которое, в частности, зависит от объемной концентрации эритроцитов в крови.
Плазменный и электростатический факторы определяют два основных механизма процесса агрегация—дезагрегация — мостиковый и электростатический. Сущность мостикового механизма заключается в том, что связующим элементом между эритроцитами в агрегате являются макро молекулярные соединения, концы молекул которых, адсорбированные на соседних клетках, образуют своеобразные «мостики». Подтверждением существования мостикового механизма является то, что расстояния между эритроцитами в агрегатах пропорциональны длине связующих макромолекул.
Применение в качестве индукторов агрегации декстранов с различной относительной молекулярной массой приводило к тому, что по мере возрастания относительной молекулярной массы декстрана расстояние между эритроцитами в агрегатах увеличивалось, оставаясь в то же время не больше длины молекулы соответствующего декстрана [Chien S. et al., 1975]. Основным пластическим материалом для межэритроцитарных мостиков в организме являются фибриноген и грубодисперсные белковые фракции, в частности у-глобулины [Лев- тов В.А. и др., 1982; Asen Р. et al., 1965].
Необходимым условием для реализации мостикового механизма является сближение эритроцитов на расстояние, не превышающее длину молекулы, образующей мостик. Увеличение концентрации эритроцитов способствует сближению клеток, а наличие сил электростатического отталкивания, создаваемых так называемым дзета-потенциалом эритроцитов, препятствует ему. Уменьшение же дзета-потенциала в свою очередь ведет к ослаблению взаимного отталкивания одноименно заряженных частиц — эритроцитов и, следовательно, способствует сближению и агрегации клеток. Это влияние заряда проявляется при разных градиентах скорости неодинаково.
Исследование реологических свойств суспензий эритроцитов в различных средах при градиентах скорости 400—800 с'1 показало, что при условиях, близких к существующим in vivo, дзета-потенциал не оказывает существенного влияния на их вязкость в этой области градиентов скорости [Сох Н., Su Goug-len, 1965]. При ацидозе, накоплении лактата, истощении щелочных резервов крови дзета-потенциал эритроцитов уменьшается, а способность клеток к склеиванию увеличивается [La Cour G. et al., 1970]. Большое значение для поверхностного заряда эритроцитов имеют сиаловые кислоты. Изучение действия нейраминидазы на эритроциты людей и животных позволило установить, что фактором, определяющим наличие у эритроцитов дзета-потенциала, являются карбоксильные группы сиаловых кислот [Cook М. et al., 1961; Eylear Е. et al., 1962].
Мостиковый и электростатический механизмы конкурируют между собой. Мощность первого из них определяется степенью связи, обеспечиваемой каждым из «мостиков», и общим количеством их. Электростатическая же сила отталкивания экспоненциально уменьшается с увеличением отношения межклеточного расстояния к толщине двойного электрического слоя вокруг эритроцита [Chien S. et al., 1976].
Механизм фиксации на эритроцитах отрицательно заряженных макромолекул: фибриногена, у-глобулинов и электрически нейтральных молекул полимеров (декстранов) пока не вполне ясен. Существует точка зрения, что сцепление молекул происходит за счет слабых водородных связей и дисперсных сил Ван-дер-Ваальса [Chien S., 1975].
Направление процесса агрегация—дезагрегация определяется результатом совокупного взаимодействия перечисленных механизмов.
Установлено, что изменение формы эритроцитов, в частности трансформация их в шиловидные или клетки с фестончатыми краями, также влияет на агрегацию [Селезнев С.А. и др., 1976].
В результате действия рассмотренных механизмов первоначально образуются двумерные эритроцитарные структуры. Затем «цепочки» эритроцитов могут соединяться. Высказывается мнение, что тип соединения зависит от связующего агента. Так, фибриноген вызывает укрупнение агрегатов, соединяя концы «цепочек», а а2-макроглобулины — соединяя их боковые поверхности [Schmid-Schonbein Н. et al., 1977]. Постепенное удлинение или ветвление агрегата запускает в действие конформационный фактор, и эритроцитарные агрегаты образуют уже трехмерную пространственную структуру. В.А. Левтов и соавт. (1982) полагают, что переход агрегатов из двух- в трехмерные является чисто количественным эффектом, физически неизбежным при достаточном их укрупнении, чем бы оно ни было вызвано.
Уменьшение ионной силы раствора усиливает дезагрегацию. Между тем если обработать эритроциты нейраминидазой, нивелирующей дзета-потенциал, торможения агрегации не наступает. Это свидетельствует о том, что действие ионной силы плазмы на ход процесса агрегация—дезагрегация опосредуется через электростатический механизм [Chien S. et al., 1976].
На процесс агрегация—дезагрегация оказывают влияние осмолярность плазмы и другие факторы, действие которых опосредовано главным образом через изменение деформируемости эритроцитов.
Важность свойств эритроцитов для процесса агрегации может быть подтверждено установлением двух основных механизмов действия антиагрегантов: они изменяют либо конформацию мембраны эритроцитов, либо, накапливаясь на их поверхности, — ее свойства [Лакин К.М., Овнатанова М.С., 1977].
Основные механизмы процесса агрегация—дезагрегация представлены на схеме 10.3.
Схема 10.3. ФАКТОРЫ, ОТРАЖАЮЩИЕ АГРЕГАЦИЮ ЭРИТРОЦИТОВ
Механизмы возникновения и развития агрегации эритроцитов весьма сложны и многообразны. Они до сих пор уточняются. Вместе с тем среди этих механизмов уже теперь можно выделить те, которые имеют ведущее значение. Заслуга детального изучения феномена внутрисосудистой агрегации эритроцитов принадлежит М. Knisely (1947, 1965). Обширный фактический материал собственных исследований и анализ работ других авторов позволили ему утверждать, что в норме ни у людей, ни у животных этот феномен практически не обнаруживается (подразумевается прижизненная агрегация эритроцитов).
Агрегаты эритроцитов при их образовании в патологических условиях закупоривают мелкие сосуды, ухудшают нутритивный кровоток и, таким образом, неблагоприятно влияют на транскапиллярный обмен [Knisely М. et al., 1947]. Крайнюю степень агрегации эритроцитов принято обозначать термином «сладжинг» (sludging). Необходимо различать агрегацию эритроцитов и их агглютинацию. Агрегация — процесс обратимый, тогда как агглютинация всегда необратима и обусловлена обычно иммунными факторами.
Результаты исследований обсуждаемого феномена с использованием метода реоскопии послужили основанием для деления эритроцитарных агрегатов на патологические и физиологические [Goldstone Y. et al., 1970]. Было установлено, что патологические агрегаты резистентны к сдвигу и не распадаются, как физиологические, а, напротив, уплотняются. Их форма и ригидность также значительно отличаются от этих параметров физиологических агрегатов. Патологические агрегаты обычно очень быстро оседают.
Выделение понятия «физиологические агрегаты» не противоречит положению об отсутствии агрегации эритроцитов в норме, а лишь дает основание говорить о едином динамическом процессе агрегация—дезагрегация, который постоянно протекает в крови. В норме дезагрегация доминирует над агрегацией.
Установлено, что интенсивность процесса дезагрегации зависит от скорости деформации. При увеличении градиента скорости число агрегатов в крови, сначала крупных, а затем и мелких (не более 30 клеток в одном агрегате), постепенно уменьшается, а при критической скорости деформации наступает полная дезагрегация [Левтов В.А. и др., 1982; Schmid-Schonbein Н. et al., 1977]. Дезагрегация сопровождается увеличением текучести крови. Полное разрушение агрегатов наступает обычно при градиентах скорости 50—80 с-1. Это свидетельствует о преобладании при этих скоростях деформации гидродинамических сил потока, стремящихся разобщить эритроциты, над силами межэритроцитарного взаимодействия. Эритроциты при этом переориентируются в потоке, стремясь обеспечить минимум диссипации энергии при наибольшей «устойчивости» [Шадрина Н.Х., 1976; Chien S., 1970].
Агрегация эритроцитов — один из важных факторов, обусловливающих нелинейность кривой течения крови. При низких скоростях деформации вклад агрегации в абсолютные значения эффективной вязкости максимален.
Результирующая направления процесса агрегация—дезагрегация в организме определяется взаимодействием по меньшей мере пяти факторов: гемодинамического, плазменного, электростатического, механического и конформационного.
Влияние гемодинамического фактора определяется зависимостью направления процесса агрегация—дезагрегация от напряжения сдвига и расстояния между отдельными клетками в потоке, которое, в частности, зависит от объемной концентрации эритроцитов в крови.
Плазменный и электростатический факторы определяют два основных механизма процесса агрегация—дезагрегация — мостиковый и электростатический. Сущность мостикового механизма заключается в том, что связующим элементом между эритроцитами в агрегате являются макро молекулярные соединения, концы молекул которых, адсорбированные на соседних клетках, образуют своеобразные «мостики». Подтверждением существования мостикового механизма является то, что расстояния между эритроцитами в агрегатах пропорциональны длине связующих макромолекул.
Применение в качестве индукторов агрегации декстранов с различной относительной молекулярной массой приводило к тому, что по мере возрастания относительной молекулярной массы декстрана расстояние между эритроцитами в агрегатах увеличивалось, оставаясь в то же время не больше длины молекулы соответствующего декстрана [Chien S. et al., 1975]. Основным пластическим материалом для межэритроцитарных мостиков в организме являются фибриноген и грубодисперсные белковые фракции, в частности у-глобулины [Лев- тов В.А. и др., 1982; Asen Р. et al., 1965].
Необходимым условием для реализации мостикового механизма является сближение эритроцитов на расстояние, не превышающее длину молекулы, образующей мостик. Увеличение концентрации эритроцитов способствует сближению клеток, а наличие сил электростатического отталкивания, создаваемых так называемым дзета-потенциалом эритроцитов, препятствует ему. Уменьшение же дзета-потенциала в свою очередь ведет к ослаблению взаимного отталкивания одноименно заряженных частиц — эритроцитов и, следовательно, способствует сближению и агрегации клеток. Это влияние заряда проявляется при разных градиентах скорости неодинаково.
Исследование реологических свойств суспензий эритроцитов в различных средах при градиентах скорости 400—800 с'1 показало, что при условиях, близких к существующим in vivo, дзета-потенциал не оказывает существенного влияния на их вязкость в этой области градиентов скорости [Сох Н., Su Goug-len, 1965]. При ацидозе, накоплении лактата, истощении щелочных резервов крови дзета-потенциал эритроцитов уменьшается, а способность клеток к склеиванию увеличивается [La Cour G. et al., 1970]. Большое значение для поверхностного заряда эритроцитов имеют сиаловые кислоты. Изучение действия нейраминидазы на эритроциты людей и животных позволило установить, что фактором, определяющим наличие у эритроцитов дзета-потенциала, являются карбоксильные группы сиаловых кислот [Cook М. et al., 1961; Eylear Е. et al., 1962].
Мостиковый и электростатический механизмы конкурируют между собой. Мощность первого из них определяется степенью связи, обеспечиваемой каждым из «мостиков», и общим количеством их. Электростатическая же сила отталкивания экспоненциально уменьшается с увеличением отношения межклеточного расстояния к толщине двойного электрического слоя вокруг эритроцита [Chien S. et al., 1976].
Механизм фиксации на эритроцитах отрицательно заряженных макромолекул: фибриногена, у-глобулинов и электрически нейтральных молекул полимеров (декстранов) пока не вполне ясен. Существует точка зрения, что сцепление молекул происходит за счет слабых водородных связей и дисперсных сил Ван-дер-Ваальса [Chien S., 1975].
Направление процесса агрегация—дезагрегация определяется результатом совокупного взаимодействия перечисленных механизмов.
Установлено, что изменение формы эритроцитов, в частности трансформация их в шиловидные или клетки с фестончатыми краями, также влияет на агрегацию [Селезнев С.А. и др., 1976].
В результате действия рассмотренных механизмов первоначально образуются двумерные эритроцитарные структуры. Затем «цепочки» эритроцитов могут соединяться. Высказывается мнение, что тип соединения зависит от связующего агента. Так, фибриноген вызывает укрупнение агрегатов, соединяя концы «цепочек», а а2-макроглобулины — соединяя их боковые поверхности [Schmid-Schonbein Н. et al., 1977]. Постепенное удлинение или ветвление агрегата запускает в действие конформационный фактор, и эритроцитарные агрегаты образуют уже трехмерную пространственную структуру. В.А. Левтов и соавт. (1982) полагают, что переход агрегатов из двух- в трехмерные является чисто количественным эффектом, физически неизбежным при достаточном их укрупнении, чем бы оно ни было вызвано.
Уменьшение ионной силы раствора усиливает дезагрегацию. Между тем если обработать эритроциты нейраминидазой, нивелирующей дзета-потенциал, торможения агрегации не наступает. Это свидетельствует о том, что действие ионной силы плазмы на ход процесса агрегация—дезагрегация опосредуется через электростатический механизм [Chien S. et al., 1976].
На процесс агрегация—дезагрегация оказывают влияние осмолярность плазмы и другие факторы, действие которых опосредовано главным образом через изменение деформируемости эритроцитов.
Важность свойств эритроцитов для процесса агрегации может быть подтверждено установлением двух основных механизмов действия антиагрегантов: они изменяют либо конформацию мембраны эритроцитов, либо, накапливаясь на их поверхности, — ее свойства [Лакин К.М., Овнатанова М.С., 1977].
Основные механизмы процесса агрегация—дезагрегация представлены на схеме 10.3.
Схема 10.3. ФАКТОРЫ, ОТРАЖАЮЩИЕ АГРЕГАЦИЮ ЭРИТРОЦИТОВ
Источник: Г.И. Назаренко, А.А. Кишкун, «Клиническая оценка результатов лабораторных исследований» 2000
А так же в разделе « Феномен внутрисосудистой агрегации форменных элементов крови »
- Пренатальная диагностика врожденных заболеваний
- Постнатальная диагностика врожденных заболеваний
- Г л а в а 10 РЕОЛОГИЧЕСКИЕ СВОЙСТВА КРОВИ И ИХ ЗНАЧЕНИЕ В КЛИНИЧЕСКОЙ ПРАКТИКЕ
- ОПРЕДЕЛЕНИЕ ПОНЯТИЙ И ЕДИНИЦ ИЗМЕРЕНИЯ
- ОСНОВЫ ТЕОРИИ НЕНЬЮТОНОВСКИХ ЖИДКОСТЕЙ
- КРОВЬ КАК НЕНЬЮТОНОВСКАЯ ЖИДКОСТЬ И ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ЕЕ РЕОЛОГИЧЕСКИЕ СВОЙСТВА
- КРОВООБРАЩЕНИЕ В СОСУДАХ ЗОНЫ МИКРОГЕМОЦИРКУЛЯЦИИ И ЕГО СВЯЗЬ С СИСТЕМНОЙ ГЕМОДИНАМИКОЙ
- СИСТЕМА МИКРОГЕМОЦИРКУЛЯЦИИ (СТРУКТУРА И ФУНКЦИИ)
- МЕТОДЫ ИССЛЕДОВАНИЯ РЕОЛОГИЧЕСКИХ И СУСПЕНЗИОННЫХ СВОЙСТВ КРОВИ
- Исследование процесса агрегации форменных элементов крови
- Исследование суспензионной стабильности крови
- Общие закономерности расстройств микрогемоциркуляции
- Синдром повышенной вязкости крови
- Нарушения проницаемости и транскапиллярного обмена
- МЕТОДЫ ИЗУЧЕНИЯ ПРОНИЦАЕМОСТИ СОСУДОВ