
Липиды (по сравнению с углеводами и белками) - наиболее богатые потенциальные источники энергии. При биологическом окислении 1 г жира освобождается 9,3 ккал энергии, в то время как при окислении 1 г углевода или белка выделяется только 4,1 ккал. В основе получения энергии микроорганизмами при использовании липидов лежат многообразные катаболические реакции, приводящие к полному расщеплению молекулы липида и освобождению энергии. Способность микроорганизмов использовать липиды определяется наличием у них активных липолитических ферментов - липаз.
Липазы микроорганизмов мало привлекали к себе внимание, так как жиры в силу устойчивости молекул, за редким исключением, не входят в состав питательных сред и практически не используются для культивирования микроорганизмов.
Большинство бактерий легко осуществляет гидролиз глицеринов (сложных эфиров глицерина и жирных кислот) и в то же время с большим трудом разрушают высшие жирные кислоты (стеариновую, пальмитиновую и др ). А некоторые бактерии - Bact. stearothermophyllus, Achromobacter lipoliticum - обладают сильно выраженной липолитической активностью. Они могут приводить к большим потерям в пищевой промышленности вследствие разрушения ими липидов.
Исследования, проведенные с липазами стафилококка показали, что эти ферменты весьма активны в период интенсивного размножения бактерий.
Гидролиз фосфолипидов осуществляется фосфолипазами или лецитиназами. Они расщепляют фосфолипиды типа лецитина и фосфатидилхолина. Лецитиназы представляют собой истинные токсины бактерий Cl. welchii (газовой гангрены).
Наличие лецитиназ присуще широкому кругу микроорганизмов различных систематических групп. Они обнаружены у многих почвенных бактерий, особенно споровых форм, актиномицетов. Бактерии, содержащие лецитиназы, играют большую роль в освобождении фосфора из органических соединений - нуклеиновых кислот, фитина и лецитина.
Окисление жирных кислот осуществляется с образованием ацетил-КоА, который играет важную роль в превращении липидов. Он активирует и переносит различные кислотные остатки (ацилы), участвует в превращении углеродных атомов углеводов, жирных кислот, глицерина и аминокислот. В обобщенном виде этот процесс может быть представлен следующим образом. Под действием липаз происходит гидролиз сложных липидов на составляющие их спирты и жирные кислоты. Так, жиры гидролизуются на глицерин и соответствующие жирные кислоты. В дальнейшем расщепление глицерина и жирных кислот идет разными путями и сопровождается образованием АТФ.
Глицерин при участии специфической фосфоферазы сначала фосфор ил ируется, затем окисляется через фосфоглицериновую кислоту до фосфоглицеринового альдегида. Последний превращается гликолитическим путем в пировиноградную кислоту, которая в зависимости от видовой специфичности микроорганизма
может давать начало одному из описанных выше путей энергетического обмена.
Расщепление образовавшихся при гидролизе свободных жирных кислот происходит также довольно сложным путем. Сначала кислота активируется коэнзимом А, и при участии АТФ образуется ацильное производное жирной кислоты - эфир КоА и жирной кислоты:
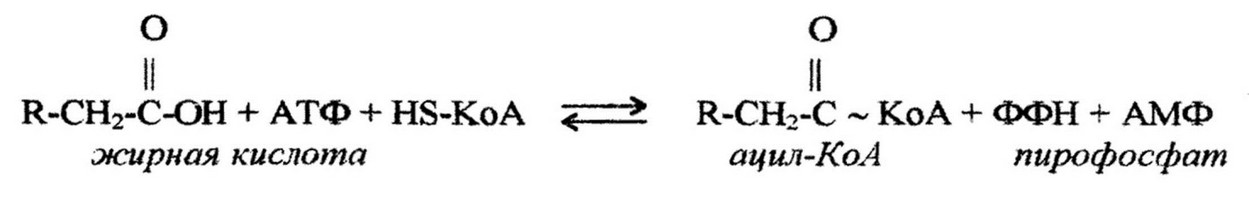
Дальнейшее превращение кислоты состоит из ряда р- окислений (всегда окисляется атом углерода, находящийся в р- положении). Сущность этого окисления состоит в следующем. Активированная жирная кислота окисляется до р-ненасыщенного производного, которое через серию реакций дегидрогенирования и гидратации превращается в
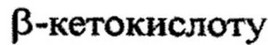 При последующем
При последующем
окислении от ,
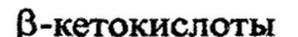 отщепляется двууглеродное
отщепляется двууглеродное
производное КоА - ацетил-КоА, на место кетонной группы присоединяется карбоксильная. Образуется новая жирная кислота, содержащая на два атома углерода меньше, чем исходная. Она подвергается таким же превращениям, как и исходная, укорачиваясь при каждом
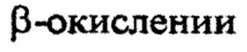 на два углеродных атома. В конечном
на два углеродных атома. В конечном
итоге вся углеродная цепочка жирной кислоты расщепляется на двууглеродные фрагменты ацетил-КоА. Последние включаются в цикл трикарбоновых кислот и окисляются до углекислоты и воды. Включение происходит путем связывания ацетильной части СН3СО со щавелево-уксусной кислотой при образовании лимонной кислоты. КоА освобождается и опять соединяется с новыми молекулами жирной кислоты.
В результате расщепления жирных кислот через ацетил-КоА высвобождается значительное количество энергии, которая запасается в АТФ. Установлено, что при полном расщеплении высокомолекулярной жирной кислоты, например, пальмитиновой, образуется 136 молекул АТФ.
В связи с тем, что липиды неодинаковы по химическому составу, пути расщепления их также различны. Но каждый из них приводит к высвобождению энергии.
Двууглеродные соединения как источник энергии. В качестве источников энергии микроорганизмы могут использовать различные двууглеродные соединения, такие как уксусная и гликолевая кислоты, этиловый спирт и др. Активированная уксусная кислота в форме ацетил-КоА может непосредственно включаться в цикл трикарбоновых кислот и полностью окисляться. Гликолевая кислота и этиловый спирт предварительно окисляются до глиоксиловой кислоты, которая в свою очередь окисляется в цикле дикарбоновых кислот. Таким же путем некоторые микроорганизмы производят и окисление ацетата. Так как этот путь обязательно включает глиоксиловую кислоту, он получил название глиоксилатного цикла (рис.7.6). Для функционирования этого цикла необходимо наличие двух дополнительных ферментов - изоцитратазы и малатсинтазы. Изоцитратаза катализирует расщепление изоцитрата с образованием глиоксиловой кислоты,
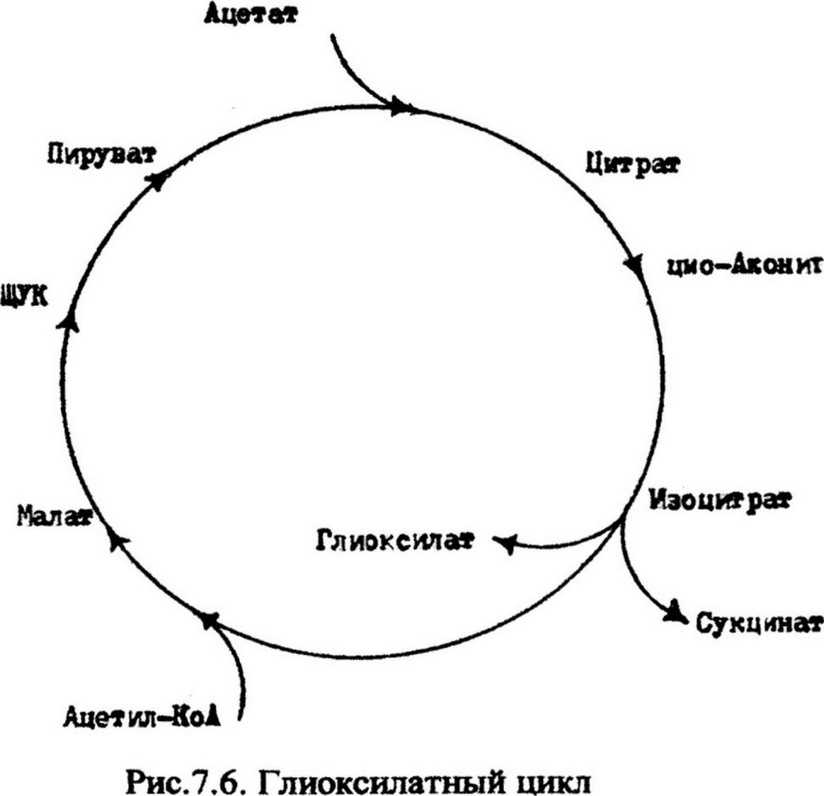
малатсинтаза - реакцию соединения глиоксиловой кислоты с ацетил-КоА в цикле дикарбоновых кислот. В результате этой реакции образуется яблочная кислота, которая затем окисляется в щавелево-уксусную и через фосфоенолпируват до пирувата.
Окисление глиоксиловой кислоты сопровождается образованием АТФ. Положительная роль цикла глиоксиловой кислоты состоит не только в образовании АТФ, но и в синтезе четырехуглеродных дикарбоновых кислот - янтарной, яблочной, щавелево-уксусной.
У бактерий рода Pseudomonas и Е. coli установлено наличие фермента изоцитратсинтазы. Он катализирует расщепление изоцитрата с образованием глиоксилата и сукцината (янтарной кислоты). Сукцинат используется микроорганизмами для синтеза щавелевоуксусной (ЩУК) кислоты, что обеспечивает непрерывность реакций цикла трикарбоновых кислот. Глиоксилатный цикл восполняет промежуточные соединения цикла Кребса, потребляемые для биосинтеза. Поэтому этот цикл называется восполняющим или анапл еретическим.