
Жизнь микроорганизмов, как и всех других организмов, связана с непрерывным расходованием энергии. Энергия необходима для переноса веществ в клетку, для осуществления движения и главным образом для синтеза органических веществ. Все реакции биосинтеза эндотермические, т. е. осуществляются с потреблением энергии. Поэтому, чтобы клетка могла синтезировать необходимые компоненты, реакции анаболизма должны быть сопряжены (связаны) с реакциями катаболизма, при которых выделяется энергия. Связь осуществляется посредством образования при катаболизме таких соединений, как АТФ (аденозинтрифосфат), УТФ (уридинтрифосфат), ЦТФ (цитидинтрифосфат), ацетил- коэнзим А (ацетил-КоА) - активированная уксусная кислота.
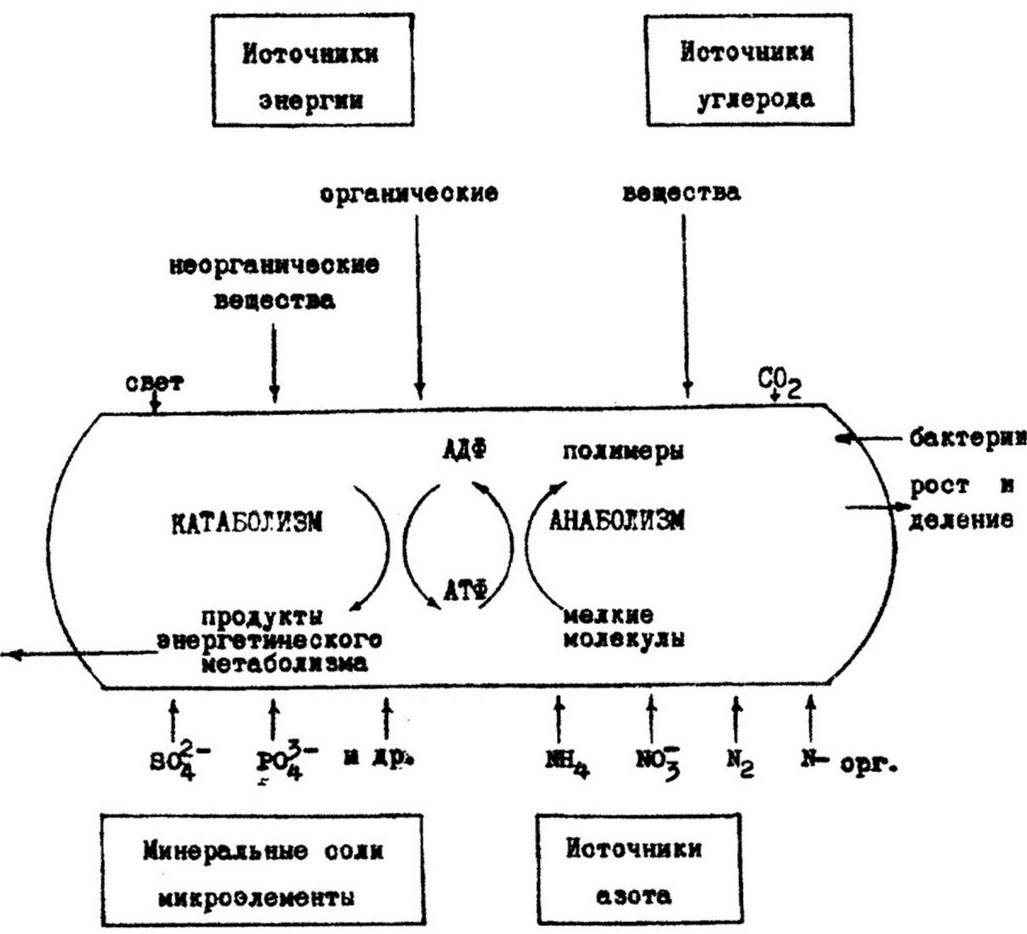
Рис. 7.1. Схема метаболизма бактерий
Центральное место в энергетическом метаболизме занимает АТФ, которая является основным донором свободной энергии в биологических системах. Это соединение легко отдает энергию и быстро регенерирует, причем отдает ее не всю сразу, а порциями. Поэтому АТФ была названа «разменной валютой» энергетического метаболизма.
Как видно из представленной формулы, АТФ - соединение нуклеотидного типа, содержащее три остатка фосфорной кислоты. Причем два концевых остатка содержат макроэргические связи, каждая из которых заключает - 34,5 кДж/моль, тогда как энергия обычной фосфатной связи составляет только - 9,6 кДж/моль.
В 60-х годах XX в. английским биохимиком Питером Митчеллом была постулирована, а потом доказана хемиосмотическая гипотеза, согласно которой в клетках существует вторая универсальная форма свободной клеточной энергии - электрохимическая, или энергия электрохимического трансмембранного градиента ионов водорода, обозначаемого
символом Л// + и измеряемого в вольтах (В, мВ).
н
Электрохимическая энергия образуется в результате работы электротранспортной цепи, функционирующей при дыхании и фотосинтезе. При транспорте электронов на цитоплазматической мембране создается трансмембранный градиент протонов водорода в силу неравномерного накопления их по обе стороны мембраны - наружной и внутренней. На цитоплазматической мембране при
функциолнировании дыхательной цепи А// + достигает 230 мВ.
н
Энергия трансмембранного потенциала первоначально появляется в электрохимической форме, затем при участии Независимого АТФ- синтетазного ферментного комплекса может превращаться в энергию АТФ. Этот ферментный комплекс локализован в цитоплазматической мембране и обеспечивает взаимное
превращение двух форм клеточной энергии (А// + АТФ).
н
Химическая энергия АТФ используется преимущественно в реакциях биосинтеза, протекающих в цитоплазме клетки.
Электрохимическая энергия трансмембранного потенциала (А/^н+)
обеспечивает процессы, локализованные только на мембране: поступление веществ в клетку, удаление продуктов метаболизма, обратный транспорт электронов с восстановлением НАД (Ф)+, работу двигательного аппарата клетки (у подвижных бактерий). Нормальная жизнедеятельность бактерий, как и других организмов, обеспечивается наличием в клетке энергии в той и другой легко мобилизируемой форме.
Большинство микроорганизмов получают энергию при расщеплении различных органических веществ - углеводов, аминокислот, липидов и других, потребляемых из внешней среды. В клетке эти вещества подвергаются окислению через ряд последовательных ферментативных реакций. В основе их лежит перенос электронов. Реакции окисления сопровождаются выделением энергии и образованием более простых соединений,
служащих исходным материалом для процессов биосинтеза. Хемолитотрофные бактерии получают энергию путем окисления неорганических соединений, таких как сероводород, аммиак, нитрит, молекулярный водород и др. Окисление осуществляется в процессе аэробного или анаэробного дыхания. В дыхательной цепи путем окислительного фосфорилирования образуется АТФ.
Углеводы как источник энергии. Для большинства гетеротрофов основным источником энергии являются углеводы Расщепление углеводов микроорганизмы производят различными путями, в которых важнейшим промежуточным продуктом, занимающим ключевую позицию, является пировиноградная кислота (пируват).
К настоящему времени у микроорганизмов хорошо изучено три основных пути расщепления углеводов: гексозодифосфатный, пентозофосфатный и путь Энтнера - Дудорова.
Гексозодифосфатный путь (гликолиз), или путь Эмбдена- Мейергофа-Парнаса (ЭМП) является главным путем, по которому расщепляет углеводы большинство микроорганизмов. Превращение углеводов гликолитическим путем начинается с фосфорилирования. При участии фермента гексокиназы и АТФ глюкоза фосфор ил ируется по шестому углеродному атому с образованием глюкозо-6-фосфата. Это активная форма глюкозы. Она служит исходным продуктом при расщеплении углеводов любым из трех указанных путей.
При гликолизе глюкозо-6-фосфат изомеризуется в фруктозо-6- фосфат, затем под действием фруктокиназы фосфор ил ируется по первому углеродному атому. Образовавшийся дифосфатный эфир сахара под действием фермента альдолазы легко распадается на две триозы: фосфоглицериновый альдегид и диоксиацетонфосфат.
Реакции между образовавшимися соединениями обратимы: диоксиацетонфосфат под действием изомеразы превращается в фосфоглицериновый альдегид. Фосфоглицериновый альдегид присоединяет остаток фосфорной кислоты и превращается в дифосфоглицериновый альдегид:
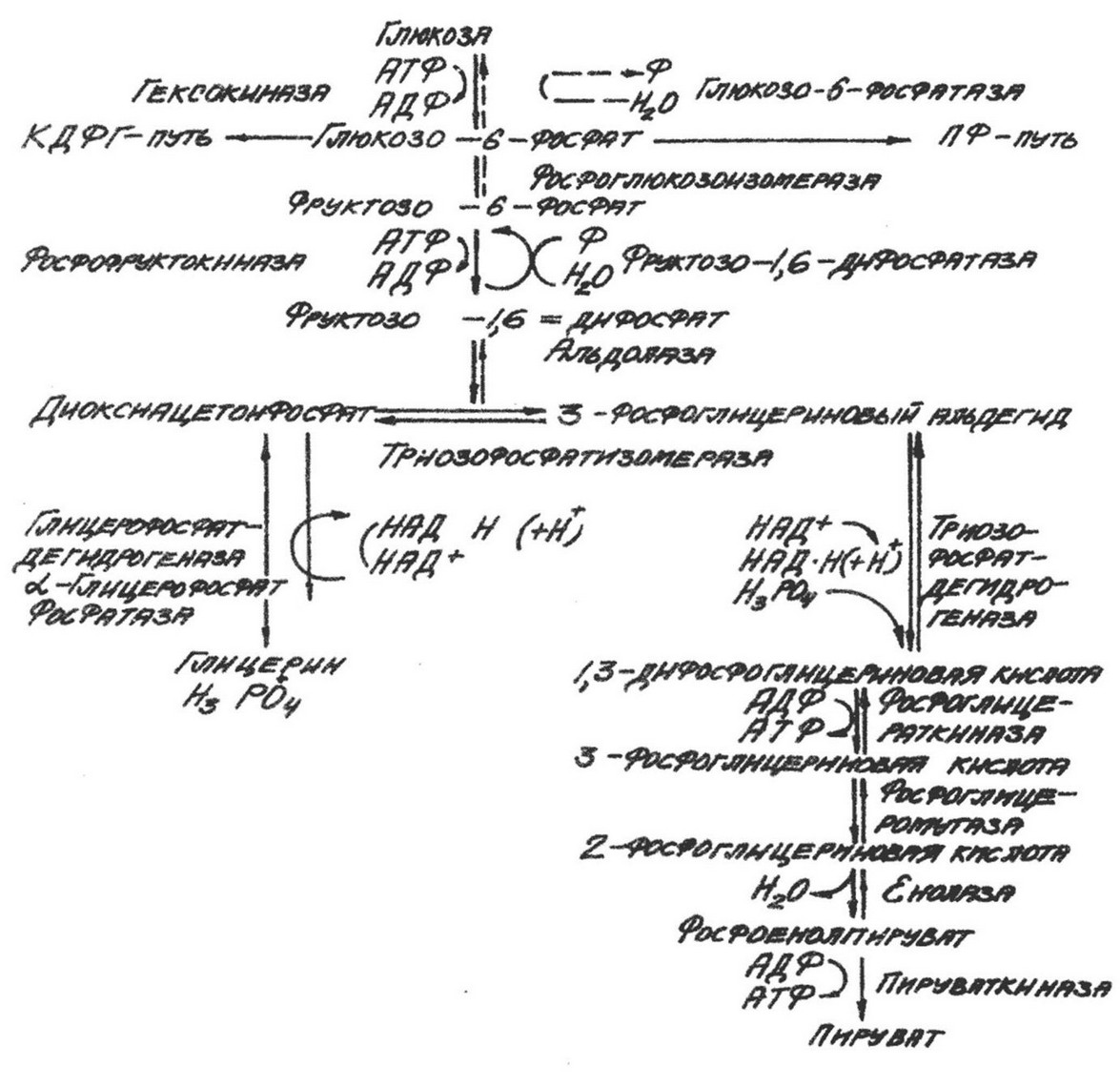
В дальнейшем происходит дегидрирование образовавшегося альдегида НАД-содержащей дегидрогеназой, в результате, чего образуется 1 -3-дифосфоглицериновая кислота и освобождается энергия, используемая клеткой для синтеза АТФ. При участии фосфоглицераткиназы богатая энергией фосфатная группа переносится на АДФ, образуется 3-фосфоглицериновая кислота и АТФ. Происходит фосфорилирование на уровне субстрата (фосфатная группа от одного субстрата передается на другой). 3-фосфоглицериновая кислота под действием фермента фосфоглицеромутазы превращается в 2-фосфоглицериновую кислоту. Фермент энолаза превращает 2-фосфоглицериновую кислоту в 2-фосфопировиноградную кислоту (энольную форму). На последнем этапе молекула фосфопировиноградной кислоты расщепляется с образованием пировиноградной кислоты, фосфат присоединяется к АДФ, превращая его в АТФ. На стадии образования пировиноградной кислоты заканчивается анаэробная фаза превращения углеводов, называемая гликолитической. В этом процессе образуется всего 2 молекулы пиру вата, 4 молекулы АТФ и 2 молекулы НАД * Н (+ tT). Но так как две молекулы АТФ первоначально затрачиваются на фосфорилирование гексозы, общий выход АТФ при превращении одной молекулы глюкозы составляет 2 молекулы. Максимальное количество энергии, получаемое клеткой при окислении одной молекулы углеводов гликолитическим путем, равно 47000 кал.
Пентозофосфатный путь (путь Варбурга-Дикенса-Хорекера) окисления углеводов широко распространен у многих организмов, но у микроорганизмов он был открыт только в 1940-1950 гг. Первоначально этот путь превращения углеводов был установлен Варбургом и Кристианом у эритроцитов и дрожжей, затем оказалось, что он характерен для всех видов семейства Enterobacteriaceae и других бактерий.
Окисление гексоз пентозофосфатным путем состоит из серии реакций, каждая из которых катализируется последовательным действием ряда ферментов. Конечными продуктами этого процесса являются 3-фосфоглицериновый альдегид и рибозо-5-фосфат.
Пентозофосфатный путь расщепления глюкозы может быть представлен следующим образом (рис.7.2). Исходный продукт глюкозо-6-фосфат дегидрируется с образованием фосфо-
глюконолактона, который затем гидролизуется до 6-фосфо- глюконовой кислоты. Фосфоглюконовая кислота в дальнейшем окисляется путем декарбоксилирования через рибулозо-5-фосфат с образованием либо рибозо-5-фосфата, либо ксилулозо-5-фосфата. которые могут превращаться в глицериновый альдегид. Последний гликолитическим путем превращается в пируват и гексозофосфаты, снова включаются в цикл. Образование рибулозы и рибозы путем окислительного декарбоксилирования выявлено у многих
микроорганизмов.
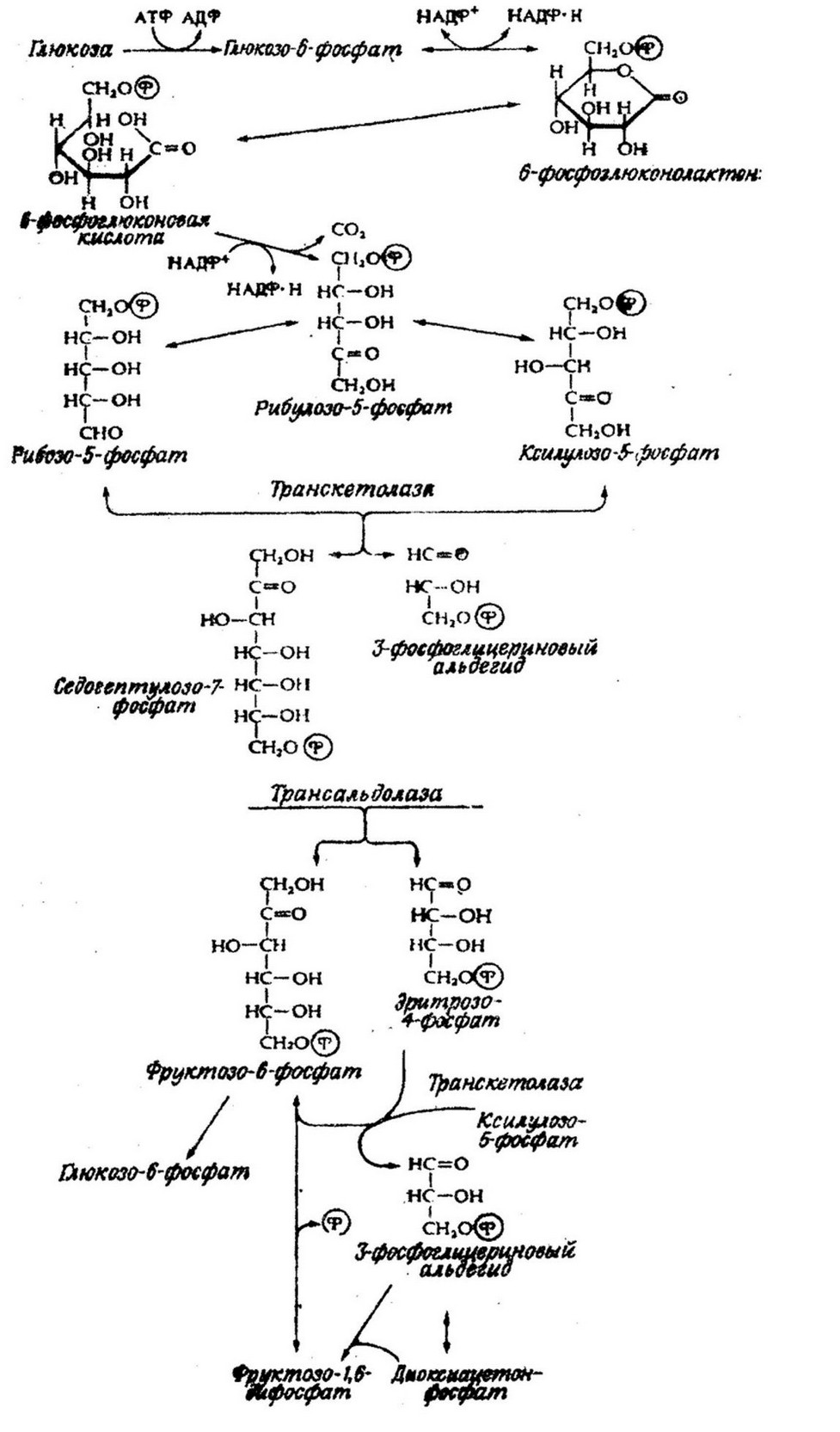
Значение пентозофосфатного пути состоит в обеспечении клетки исходными веществами для биосинтеза. Образующиеся пентозофосфаты являются предшественниками нуклеотидов и нуклеиновых кислот. Кроме того, важная функция пентозофосфатного пути заключается в снабжении клетки НАДФ * Н, необходимым для осуществления восстановительных реакций биосинтеза. Как механизм получения энергии этот путь в два раза менее эффективен, чем гликолитический: на каждую молекулу глюкозы образуется молекула АТФ. Он рассматривается как дополнительный путь окисления углеводов. Это как бы «шунт» гликолитического пути.
Путь Энтнера-Дудорова (КДФГ-путь) открыт у
микроорганизмов в 1952 г. Он менее распространен, чем два предыдущих и возможен лишь у некоторых групп прокариот. По этому пути окисление углеводов производят энтеробактерии, азотобактер, клубеньковые бактерии, отдельные псевдомонады и уксуснокислые бактерии.
Первым продуктом данного пути является глюкозо-6-фосфат. Она дегидрируется до 6-фосфоглюконовой кислоты. Затем после дегидратации образуется продукт, характерный только для пути Энтнера-Дудорова - 2-кето-3-дезокси-6-фосфоглюконовая кислота (КДФГК). Последняя расщепляется на пируват и 3-фосфо- глицериновый альдегид, который под действием ферментов гликолиза превращается во вторую молекулу пировиноградной кислоты:
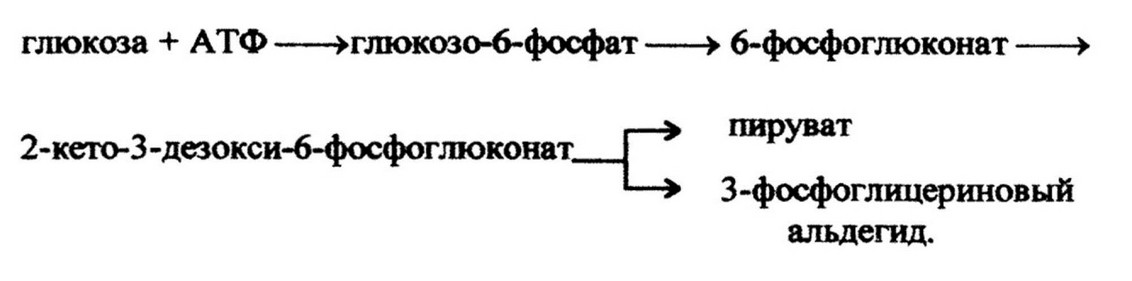
При окислении одной молекулы глюкозы КДФГ -путем образуется одна молекула АТФ и две молекулы НАДФ' Н. Этот путь обеспечивает использование микроорганизмами глюконовой кислоты.
Таким образом, основным ключевым продуктом расщепления углеводов разными путями является пировиноградная кислота, которая при участии ферментов превращается в различные интермедиаты.
Полное окисление пировиноградной кислоты происходит в цикле трикарбоновых кислот (цикл Кребса) и дыхательной цепи.
Цикл трикарбоновых кислот. Включению пировиноградной кислоты в цикл Кребса предшествует окисление ее до ацетил- коэнзима А (КоА). Это осуществляется в результате сложной реакции, состоящей из окислительного декарбоксилирования пирувата и активирования образовавшегося ацетата при посредстве КоА. Окисление пировиноградной кислоты до ацетил-КоА катализируется пируватдегидрогеназной системой:
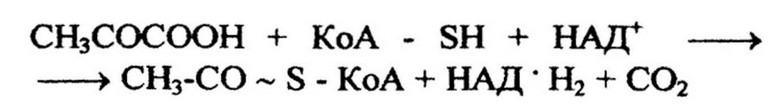
Ацетил-КоА под действием цитрат-синтазы вступает в реакцию со щавелевоуксусной кислотой, образуя лимонную кислоту, которая является основным звеном цикла трикарбоновых кислот (рис.7.3). Лимонная кислота претерпевает ряд превращений и в результате образуется а-оксоглутаровая кислота. Реакция дегидрирования изолимонной кислоты сопряжена с восстановлением одной молекулы НАД до НАДН. а-оксоглутаровая кислота в свою оцередь подвергается окислительному декарбоксилированию с образованием сукцинил-КоА. Это
соединение содержит высокоэнергетическую тиоэфирную связь. На следующем этапе цикла тиоэфирная связь сукцинил-КоА разрывается, образуется янтарная кислота (сукцинат), а
освобождающаяся энергия используется для синтеза АТФ из АДФ и неорганического фосфата ( у животных фосфорилируется не АТФ, а ГТФ). Следовательно, в данном случае имеет место субстратное фосфорилирование.
Янтарная кислота окисляется в дальнейшем сукцинатдегидрогеназой до фумаровой кислоты. Последняя под действием фумаразы гидратируется с образованием яблочной кислоты, котора посредством дегидрирования превращается в
щавелевоуксусную кислоту. Эта реакция завершает полный оборот цикла трикарбоновых кислот. Затем происходит конденсация образовавшейся молекулы щавелевоуксусной кислоты с другой молекулой ацетил-КоА и запускается следующий оборот цикла.
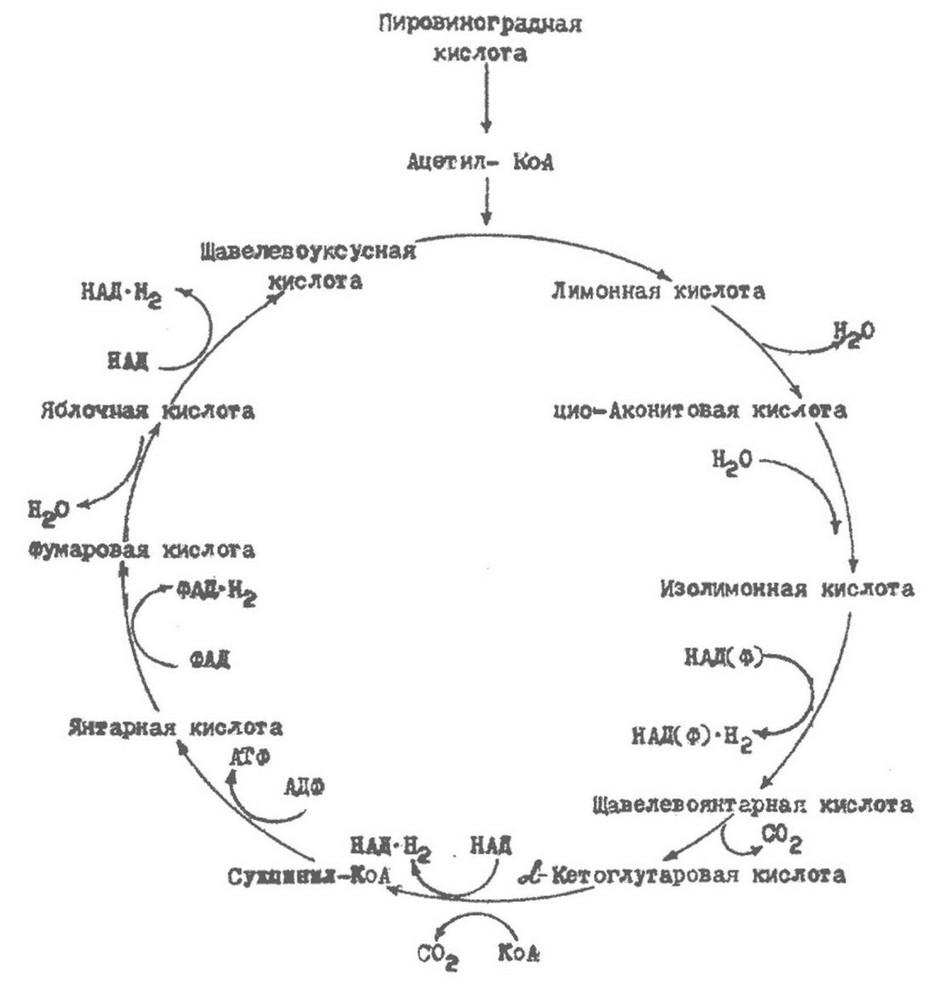
Рис. 7.3. Цикл трикарбоновых кислот
В результате одного оборота цикла образуется 3 молекулы НАД ’ Н2 и 1 молекула АТФ.
Цикл трикарбоновых кислот обеспечивает полное окисление пировиноградной кислоты до С02 и Н20. Кроме того, в цикле трикарбоновых кислот образуется ряд интермедиатов, как а- оксоглутаровая, щавелевоуксусная, янтарная, фумаровая кислоты, которые необходимы для биосинтетических процессов. Поэтому не
только аэробы, а даже строгие анаэробы содержат многие ферменты этого цикла.
Таким образом, основными метаболическими путями, присущими почти всем микроорганизмам, являются: гликолиз, который протекает в цитозоле и выполняет две важные функции - расщепляет глюкозу с образованием АТФ и поставляет углеродные скелеты молекул для биосинтеза; пентозофосфатный путь, протекающий также в цитозоле и выполняющий две основные функции - генерирование НАДФ * Н для восстановительных реакций биосинтеза и образование рибозо-5-фосфата для синтеза нуклеотидов; цикл трикарбоновых кислот, функционирующий исключительно в мембранах. Он осуществляет заключительные этапы окисления источников энергии - углеводов, аминокислот, органических кислот.