
Дыхание - метаболический процесс протекающий в клетках с освобождением энергии и генерированием АТФ, в котором конечным акцептором электронов (водорода) служат неорганические соединения. В зависимости от конечного акцептора электронов различают аэробное и анаэробное дыхание. При аэробном дыхании акцептором водорода является кислород, при анаэробном - неорганические окисленные соединения типа нитратов и сульфатов.
Аэробное дыхание. В качестве энергетического субстрата для дыхательного метаболизма микроорганизмы используют широкий круг природных соединений. Независимо от сложности структуры окисляемого субстрата потребление его в качестве источника энергии основано на одном и том же принципе: постепенное расщепление до образования простых соединений, способных вступать в реакции цикла трикарбоновых кислот. Таким соединением основных метаболических путей является пируват.
Окисление пирувата при аэробном дыхании осуществляется в цикле Кребса, в который он поступает при посредстве ацетил-КоА. Полное окисление его приводит к освобождению двух молекул углекислоты и восьми атомов водорода. Акцептором водорода, как указано выше, у аэробных бактерий является кислород. Передача
водорода (электронов) на кислород осуществляется через последовательную цепь молекул-переносчиков, так называемую дыхательную цепь, или цепь транспорта электронов:
Субстрат gt; НАД* Н2 gt; Флавопротеид gt; Кофермент О
gt; Цитохром с gt; Цитохром аа3 gt; 02
Дыхательная цепь представляет собой систему пространственно организованных молекул-переносчиков, осуществляющих перенос электронов от окисляемого субстрата к акцептору. Она развита у аэробов и факультативных анаэробов, только у последних терминальным акцептором электронов, кроме кислорода, являются нитраты и сульфаты.
Компонентами дыхательной цепи, локализованными в мембране, являются такие переносчики белковой природы, как флавопротеиды, FeS-белки, цитохромы, и небелковой природы - хиноны (убихиноны, менахиноны). НАД(Ф)-зависимые дегидрогеназы, отщепляющие водород от окисляемого субстрата - растворимые ферменты; флавопротеидные дегидрогеназы могут находиться либо в мембране, либо в растворимой форме в цитоплазме.
 хиноны осуществляют перенос атомов и цитохромы - электронов. Так как содержащие переносчики электронов, погружены в цитоплазму, то имеется прямое взаимодействие между дыхательной цепью, с одной стороны, и окисляемым субстратом, АДФ и неорганическим фосфатом цитоплазмы - с другой. Такое свободное взаимодействие дыхательной цепи с цитоплазмой определяет отличительные особенности функционирования дыхательного аппарата прокариот от эукариот. Так, дыхательные цепи прокариот менее стабильны по сотаву переносчиков электронов и энергетически менее эффективны. В дыхательной цепи эукариот имеются три участка, где происходит выброс протонов и
хиноны осуществляют перенос атомов и цитохромы - электронов. Так как содержащие переносчики электронов, погружены в цитоплазму, то имеется прямое взаимодействие между дыхательной цепью, с одной стороны, и окисляемым субстратом, АДФ и неорганическим фосфатом цитоплазмы - с другой. Такое свободное взаимодействие дыхательной цепи с цитоплазмой определяет отличительные особенности функционирования дыхательного аппарата прокариот от эукариот. Так, дыхательные цепи прокариот менее стабильны по сотаву переносчиков электронов и энергетически менее эффективны. В дыхательной цепи эукариот имеются три участка, где происходит выброс протонов и
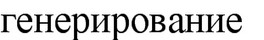 А/л + , у большинства прокариот - только один- н
А/л + , у большинства прокариот - только один- н
два участка, т. е. суммарный выход энергии у прокариот ниже.
Функционирование дыхательной цепи осуществляется
следующим образом. Водород окисляемого субстрата,
освобожденный в реакциях цикла Кребса или мобилизованный непосредственно НАД (Н2)-зависимыми дегидрогеназами передается в дыхательную цепь на флавиновые дегидрогеназы, затем на убихиноны. Здесь атом водорода расщепляется на протон и электрон. Протон выделяется в среду, электрон передается на систему цитохромов до цитохромоксидазы. Она передает электрон на кислород-терминальный акцептор, который активизируется и соединяется с водородом, образуя воду и перекиси. Последние разлагаются каталазой на воду и кислород. Перенос электронов приводит к значительному изменению свободной энергии в клетке.
Расчеты энергетического баланса показали, что при расщеплении глюкозы гликолитическим путем и через цикл традсарбоновых кислот с последующим окислением в дыхательной цепи до С02 и Н20 на каждый моль глюкозы образуется 38 молей АТФ. Причем максимальное количество АТФ образуется в дыхательной цепи - 34 моля; 2 моля - в ЭПМ-пути и 2 - в ЦТК.
Ввиду большого разнообразия ферментных систем, входящих в дыхательную цепь, окисляемых субстратов и терминальных акцепторов у бактерий существует большое количество разнообразных дыхательных цепей. Так, в дыхательной цепи уксусно-кислых бактерий отсутствуют цитохромы а + аз: дегидрогеназы -*С -gt;Cj -gt;Aj -gt;02. Еще меньший набор компонентов имеет дыхательная цепь Agrobacterium tumefaciens: НАДН дегидрогеназа -»Q —»С —Ю2. Дыхательная цепь клубеньковых бактерий и азотобактера характеризуется наличием разнообразных цитохромов: дегидрогеназы —gt; Ь—gt; с—> а—gt; аз —gt;02. Укороченные дыхательные цепи характерны для многих бактерий. В энергетическом обмене они менее полезны для бактерий из-за низкого выхода АТФ.
Биолюминесценция. У некоторых бактерий существует ответвление от основнорй дыхательной цепи. Электроны от НАД передаются не на ФАД а на ФМН (флавомононуклеотид). Последний реагирует с ферментом люциферазой, кислородом и альдегидом пальмитиновой кислоты. Люцифераза (Л) катализирует
реакцию восстановительного альдегида (АН2) с АТФ (продукт этой реакции при последующем окислении испускает видимый свет):

Эта реакция получила название «светлячковой» из-за ее наличия у светлячка Photinus piralis. Ее используют для количественного определения АТФ, потому что интенсивность свечения находится в прямой зависимости от количества АТФ.
Механизм биолюминесценции состоит в том, что в результате взаимодействия ФМН с люциферазой, кислородом и альдегидом электроны в некоторых молекулах переходят в возбужденное состояние и возвращение их на основной уровень сопровождается испусканием света. Образования АТФ при люминесценции не происходит. Поэтому эффективность функционирования дыхательной цепи снижается, т. е. клетка не получает всей энергии, заключенной в окисляемом субстрате, так как часть ее превращается в световую.
Свечение бывает тем интенсивнее, чем лучше условия аэрации культуры. Светящиеся бактерии являются весьма чувствительными индикаторами молекулярного кислорода. М. Бейеринк применял светящиеся бактерии в качестве индикатора для обнаружения кислорода при бактериальном фотосинтезе (в те времена не было известно, что бактериальный фотосинтез протекает без выделения кислорода).
Способностью к биолюминесценции обладают факультативно-анаэробные морские бактерии, объединенные в род Photobacterium (светящиеся бактерии).В аэробных условиях они окисляют органические субстраты с испусканием лунно-голубого света. Биолюминесценция рассматривается как приспособление некоторых микроорганизмов к защите от вредного действия кислорода.
Неполное окисление. Большинство аэробных микроорганизмов в процессе дыхания осуществляют полное окисление углеводов до углекислоты и водь*. При этом
высвобождается вся энергия, заключенная в субстрате. Примером может служить окисление глюкозы пекарскими дрожжами:
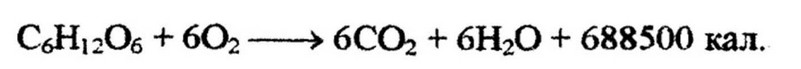
Однако окисление может быть и неполным. Это зависит от видовой принадлежности микробов и условий развития. Обычно неполное окисление наблюдается при избытке в среде углеводов и недостатке кислорода. Конечными продуктами неполного окисления являются органические кислоты, такие как уксусная, лимонная, фумаровая, глюконовая и др. Типичным примером неполного окисления является образование уксусной кислоты из спирта бактериями рода Acetobacter:
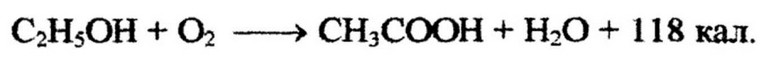
Этот окислительный процесс используется микроорганизмами для получения энергии. При неполном окислении образование макроэргических фосфатных связей происходит в процессе переноса электронов. Однако общий выход энергии при этом значительно меньший, чем при полном окислении. Часть энергии окисляемого исходного субстрата сохраняется в образующихся органических кислотах. В связи с тем, что сходные кислоты (янтарная, молочная) образуются при брожении углеводов, неполное окисление называют «окислительным брожением». Отличительной особенностью неполного окисления является участие кислорода в реакциях. Поэтому аэрация - необходимое условие образования органических кислот микроорганизмами. Установлено, что образование а- глутаминовой кислоты бактериями (Corynebacterium glutamicum) происходит только в строго аэробных условиях. Причем выход данной аминокислоты может быть очень высоким - 0,6 моля глутамина на 1 моль использованной глюкозы.
Микроорганизмы, развивающиеся за счет энергии неполного окисления, используются в микробиологической промышленности для получения органических кислот, в том числе и аминокислот.
Анаэробное дыхание. В анаэробных условиях, т. е при отсутствии молекулярного кислорода, некоторые микроорганизмы,
такие как Micrococcus denitrificans и бактерии родов Desulfovibrio и Desulfotomaculum в качестве акцептора водорода используют окисленные минеральные соединения - нитраты, сульфаты, которые легко отдают кислород, превращаясь в восстановленные формы. Продуктами восстановления нитратов служит нитрит и молекулярный азот; сульфаты восстанавливаются до сероводорода и других соединений. Образовавшиеся восстановленные продукты выделяются из клетки. Окисление органического вещества в анаэробных условиях происходит путем дегидрогенирования Отщепляемый водород поступает в дыхательную цепь и переносится на соответствующий акцептор. Конечная реакция катализируется нитратредуктазой. Последняя в анаэробных условиях функционирует как цитохромоксидаза.
Нитратредуктаза является индуцибельным ферментом. Синтез ее происходит только в анаэробных условиях при наличии нитрата. Кислород ингибирует синтез нитратредуктазы. При наличии нитратредуктазы в клетке (если бактерии из анаэробных условий переносятся в аэробные) кислород конкурирует с нитратом за электроны в дыхательной цепи, подавляя тем самым функции данного фермента. Вот почему нитратное и сульфатное дыхание осуществляется только в анаэробных условиях.
Способность микроорганизмов использовать в качестве акцепторов электронов нитраты и сульфаты позволяет производить им полное окисление субстрата и получать таким путем необходимое количество энергии. Так, денитрифицирующие бактерии при нитратном дыхании производят полное окисление органических субстратов, выход энергии при этом только на 10% ниже, чем при аэробном дыхании. Образование АТФ происходит в результате фосфор ил ирова ния в дыхательной цепи.