
Микроорганизмы в отличие от растений и животных характеризуются многообразием типов питания. Это связано с их способностью использовать разные источники энергии и разные соединения доя конструктивного метаболизма.
Источником углерода для одних микроорганизмов могут служить неорганические соединения, в частности углекислота, для других - органические вещества. Те микроорганизмы, которые получают углерод только из углекислоты и синтезируют из него необходимые органические вещества, называются автотрофами (от греч. autos - сам, trophe - питание). Они обладают наибольшей автономностью и независимостью своего развития от жизнедеятельности других организмов, могут жить в чисто минеральных средах. Другие микроорганизмы используют углерод из органических соединений и называются гетеротрофами (от греч. heteros - другой, trophe - питание). Они способны развиваться только в средах, содержащих органические соединения.
Восстановление углерода до органических соединений требует затрат энергии. Источником энергии может быть солнечный свет и химические связи органических и минеральных веществ Организмы, которые используют световую энергию, называются фототрофами, или фотосинтезирующими, а те, которые используют энергию химических реакций (химическую энергию), называются хемотрофами, или хемосинтезирующими. Соответствующие
названия имеют и микроорганизмы, обладающие этими типами питания.
К фотолитотрофам относятся цианобактерии, фотосинтезирующие бактерии, использующие для процессов синтеза энергию солнечного света. Подобно высшим растениям в цитоплазме клеток этих бактерий содержатся пигменты типа хлорофилла. Бактериальный хлорофилл по химической структуре весьма сходен с хлорофиллом растений. Основное различие между ними состоит в том, что у бактериохлорофилла в первом пирольном кольце вместо винильной группы (СН2 = СН-), характерной для хлорофилла растений, находится ацетильная группа (СН3СО-), а также содержится на два атома водорода больше. Максимумы поглощения света этих двух хлорофиллов не совпадают.
Кроме хлорофилла все фотосинтезирующие бактерии содержат каротиноиды, которые также принимают участие в фотосинтезе, передавая энергию поглощаемого света бактериохлорофиллу.
В основе бактериального фотосинтеза лежит превращение световой энергии, поглощенной пигментами, в химическую энергию макроэргических связей АТФ, образуемой в процессе фотофосфорилирования и используемой затем для усвоения углекислого газа и процессов биосинтеза.
В общих чертах процесс фотосинтеза у бактерий происходит так же, как у зеленых растений. Но у растений источником водорода является вода, которая окисляется до кислорода. У фотолитотрофных бактерий источником водорода для фотосинтеза является сероводород или молекулярный водород. Поэтому кислород никогда при этом процессе не выделяется. Еще одно отличие: растения для восстановления одной молекулы углекислоты используют четыре кванта энергии, бактерии - только один квант. Конечные продукты фотосинтеза у растений и бактерий одинаковы: это соединения типа углеводов.
Бактериальный и растительный фотосинтез может быть представлен следующими обобщенными уравнениями:
свет
С022Н2А gt; [СН20] + Н20 + 2 А ( Н2А - донор водорода)
бактериальный фотосинтез

К фотосинтезирующим бактериям относятся: зеленые
бактерии, пурпурные серобактерии и пурпурные несеробактерии.
К фотоорганотрофам относятся несерные пурпурные бактерии, которые для восстановления углекислого газа могут использовать водород органических соединений. Пурпурные несерные бактерии входят в состав сем. Rhodospirillaceae и представлены двумя родами: Rhodospirillum - клетки спиральной формы и Rhodopseudomonas - клетки палочковидной формы. Исследования физиологии данных бактерий проведены на чистых культурах Ван-Нилем. В отличие от серобактерий они являются факультативными анаэробами, нуждаются в витаминах, хорошо развиваются на органических средах. Причем развитие их может происходить как на свету, так и в темноте. Эти бактерии используют в качестве источника энергии или солнечный свет, или аэробное окисление. На свету они могут развиваться в строго анаэробных условиях. В отличие от серобактерий, которые для фотосинтеза используют водород сероводорода или других соединений серы, у пурпурных несерных бактерий донором водорода служат органические соединения. На свету акцептором водорода является углекислота. Вначале происходит окисление органического вещества путем дегидрирования, т. е. отнятием водорода, затем этот водород переносится на молекулы углекислоты:
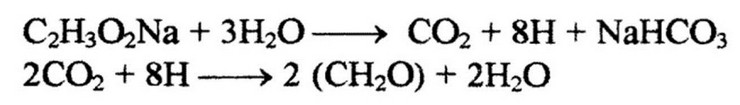
В темноте акцептором водорода в аэробных условиях является кислород, в анаэробных - сера. Поэтому развитие несерных пурпурных бактерий в темноте возможно только при наличии кислорода или серы. Эти микроорганизмы характеризуются полным набором основных дыхательных ферментов, переносчиков водорода: НАД, ФАД, цитохромы. Окисление субстрата происходит по циклу трикарбоновых кислот. Пурпурные бактерии используют
органические вещества не только в качестве доноров водорода, но и как непосредственные источники углерода.
К хемолитотрофам относятся бактерии, которые способны ассимилировать углекислоту и синтезировать органические вещества за счет химической энергии, получаемой при окислении различных минеральных веществ: аммиака, нитритов, сероводорода, водорода и железа. В отличие от фотосинтеза, где используется энергия света, этот процесс получил название хемосинтеза. Хемосинтез открыт в 1887 г. С. Н. Виноградским у бесцветных серобактерий, затем у ряда других микроорганизмов.
Большинство хемолитотрофных бактерий способно развиваться в чисто минеральных средах, используя углекислоту в качестве источника углерода, а минеральные вещества в качестве источников энергии и электронов, окисляя их в процессе дыхания. Все хемолитотрофные бактерии ассимилируют СО2 через цикл Кальвина. У облигатных хемолитотрофов (тиобацилл и нитрифицирующих бактерий) имеются специальные структуры - карбоксисомы, содержащие фермент рибулозодифосфаткарбок- силазу, с помощью которого осуществляется фиксация СО2. Неспособность облигатных хемолитотрофов использовать органические субстраты связана с тем, что у них не функционирует цикл Кребса из-за отсутствия одного из ключевых ферментов цикла
4
а-кетоглуторатдегидрогеназы.
Существует несколько физиологических групп хемолитотрофных бактерий.
Они характеризуются специфичностью в отношении окисляемого субстрата. Например, нитрифицирующие бактерии рода Nitrosomonas окисляют аммиак до нитрита, a Nitrobacter окисляет нитрит до нитрата:
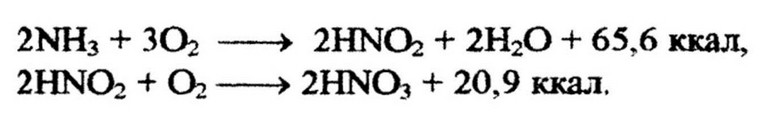
Бесцветные серобактерии окисляют сероводород до элементарной серы, а затем серу до серной кислоты:
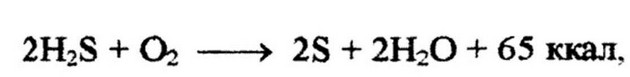
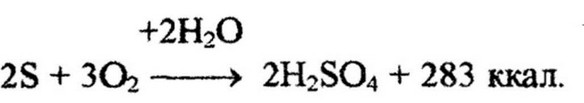
Железобактерии окисляют закисное железо в окисное:
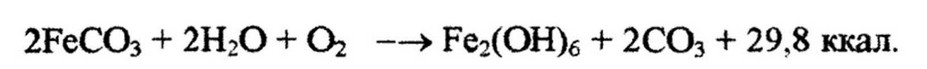
Энергия, выделяющая в результате окислительных реакций, используется хемолитотрофами для восстановления углекислоты. На восстановление молекулы углекислого газа требуется больше энергии, чем ее выделяется при окислении молекулы аммиака или сероводорода, к тому же коэффициент использования выделяющейся энергии невелик ( у нитрификаторов 5-10 %). В связи с этим хемолитотрофным микроорганизмам приходится перерабатывать очень большое количество веществ по сравнению с тем количеством органического вещества, которое они синтезируют. Например, Nitrosomonas окисляет до 35 молекул аммиака на одну молекулу восстановленной углекислоты (Работнова, 1966):
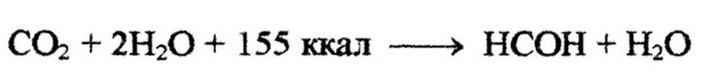
Образующийся муравьиный альдегид или другое подобное соединение полимеризуется в углеводы. Хемолитотрофные организмы, окисляющие неорганические вещества, являются важнейшими геохимическими агентами. С их деятельностью в природе связано образование и разрушение полезных ископаемых, они осуществляют важнейшие этапы круговорота минеральных элементов.
К хемоорганотрофам относятся микроорганизмы, которые нуждаются в готовых органических соединениях. Источником энергии и источником водорода для них служат органические вещества. Это самая разнообразная и весьма многочисленная группа микроорганизмов. Они широко распространены в природе и играют огромную роль в разложении органических веществ. В качестве источников углерода хемоорганотрофы используют готовые органические соединения самой различной химической структуры.
Наиболее подходящими являются соединения, содержащие альдегидные и кетонные группы, а также насыщенные связи.
Кроме органического углерода, гетеротрофные микроорганизмы нуждаются в углекислоте. Последняя играет важную роль в промежуточном обмене. Чаще всего гетеротрофы используют углекислоту для карбоксилирования трехуглеродных соединений - пирувата и фосфоенолпирувата, в результате чего образуется щавелевоуксусная кислота, легко вступающая в дальнейшее превращения, например, в цикле трикарбоновых кислот. Образующиеся промежуточные продукты используются клеткой для биосинтеза макромолекул.
Из хемоорганотрофов наибольшей степенью гетеротрофности характеризуются облигатные внутриклеточные паразиты, т. е. микроорганизмы, которые могут жить только внутри клеток других организмов, К ним относятся бактерии порядков Rickettsiales и Chlamydiales. Паразитический образ жизни привел к упрощению у них ферментных систем энергетического метаболизма. Например, у некоторых хламидий отсутствует система АТФ-АДФ, ферменты гликолиза, ЦТК. Это поставило их в полную зависимость от клеток- хозяев, неспособность к росту на питательных средах. В отличие от них, факультативные паразиты могут развиваться на искусственных питательных средах, хотя и очень сложного состава, включающего мясной автолизат, нативную кровь или сыворотку, набор витаминов и т. д.
Кроме паразитов, к гетеротрофам относится большая группа сапрофитных бактерий (от греч. sapros - гнилой, phyton - растение), которые также развиваются за счет готовых органических соединений. В отличие от паразитов, сапрофиты используют отмершие ткани растений и животных, продукты жизнедеятельности разных организмов. В. Л. Омелянский назвал эту группу бактерий «могильщиками органического мира».