
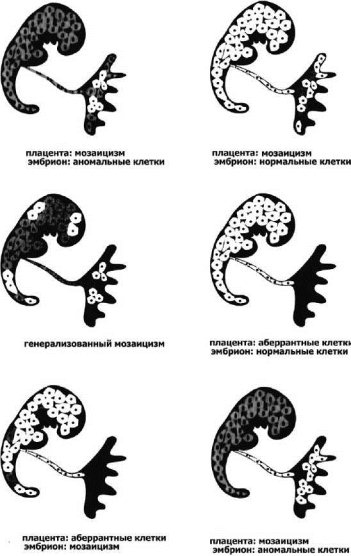
Если жизнеспособная клеточная линия с анеуплоидией возникает в процессе первых делений дробления, морула будет в большей или меньшей степени мозаичной. Очевидно, что размеры аберрантного клона будут определяться стадией возникновения мутации, а также локализацией аномальной клетки в моруле (рис. 7.2, 7.3, 7.4). Если распределение двух клеточных линий происходит случайно, возрастает вероятность возникновения истинных мозаиков с двумя линиями клеток в хорионе и в тканях плода (рис. 7.4, 7.5).
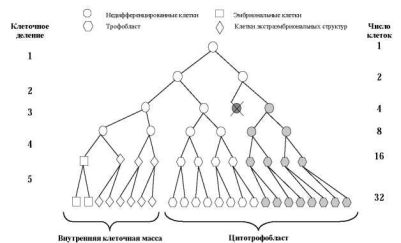
Рис. 7.2. Модель возникновения ограниченного плацентарного мозаицизма при нерасхождении хромосом во втором делении дробления
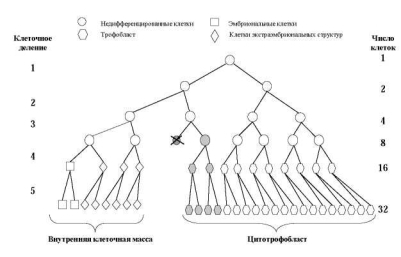
Рис. 7.3. Модель возникновения ограниченного плацентарного мозаицизма при нерасхождении хромосом в третьем делении дробления
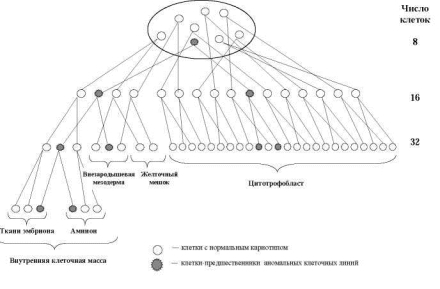
Рис. 7.4. Модель возникновения хромосомного мозаицизма в раннем эмбриогенезе человека
Как показали молекулярно-цитогенетические исследования, около 50 % дробящихся эмбрионов человека являются анеуплоидными (см. главу 5). При этом 38-46 % морфологически нормальных эмбрионов на стадии 8-10 клеток имеют три и более анеуплоидных бластомера [654]. Число бластомеров, отличающихся по набору хромосом, может достигать половины всех клеток зародыша [331, 653]. Экстраполируя результаты исследования анеуплоидии по четырем парам хромосом методом FISH (X, Y, 13, 18, 21) на весь набор, можно ожидать, что частота мозаичных эмбрионов в период дробления составит не менее 80 % [388]. Если это предположение подтвердится в дальнейших исследованиях, то хромосомный мозаицизм следует признать особенностью нормального развития зародыша человека, по крайней мере, до стадии 8-10 клеток.
Аномальная сегрегация хромосом может быть обусловлена такими факторами как репликация без цитокинеза, фрагментация ядра, аномалии движения хромосом в анафазе [654]. Критическим для нормального расхождения хромосом являются завершение репликации, расположение
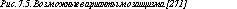
хромосом на экваторе и прикрепление кинетохор к микротрубочкам веретена. Незавершенность одного из этих событий приводит к блокированию клеточного цикла на соответствующей стадии и, в конечном счете, — к гибели клетки. Высокая частота аутосомных моносомий и трисомий у дробящихся эмбрионов, сопоставимая с частотой анеуплоидных ооцитов [654], свидетельствует об отсутствии на стадии дробления зиготы механизма селекции клеток с хромосомным дисбалансом [567]. Не исключено, что механизмы, контролирующие клеточный цикл, вступают в силу только после активации генома самого зародыша [455, 834]. По-видимому, процесс восстановления механизмов контроля митотического цикла происходит асинхронно и выраженно не во всех клетках. Так, доля гете- роплоидных бластомеров на стадии бластоцисты (~ 64 клетки), как правило, снижается по сравнению с дробящимися эмбрионами и составляет в среднем 10,5 % [414]. Однако частота морфологически нормальных эмбрионов с мозаичной формой анеуплоидии остается высокой и достигает 40 % [309]. Следовательно, мозаичные эмбрионы, имеющие подавляющее большинство гетероплоидных бластомеров, подвергаются элиминации, и переход из стадии морулы в стадию бластоцисты является критическим для дальнейшего развития эмбриона [414].