
Щитовидная железа секретирует регуляторы основного обмена: йодсодержащие гормоны — трийодтиронин (Js) и тироксин (T4), а также кальцитонин — один из эндокринных регуляторов обмена Ca2+. Йодсодержащие гормоны вырабатывают эпителиальные клетки стенки фолликулов, кальцитонин — светлые клетки. Две пары паращитовидных желёз (верхние и нижние), секретирующие антагонист кальцитонина — паратиреокрин, почти всегда анатомически тесно связаны с щитовидной железой.
А. Источники. Эндокринные клетки щитовидной железы имеют двоякое происхождение: из стенки глотки и нервного гребеня.
- Глотка. Эпителий бранхиогенной группы желёз (щитовидная, вилочковая, околощито- видные) развивается из энтодермы глоточных карманов (см. главу 12 I Б 2 а),
а. Паращитовидные железы
- Нижние — третья пара глоточных карманов.
- Верхние — четвёртая пара.
б. Щитовидная железа. Зачаток щитовидной железы в виде выпячивания крыши глотки между первой и второй парами глоточных карманов (у корня языка) возникает на 3-4-й неделе внутриутробного развития. Эпителиальный зачаток железы растёт вентральнее хрящей гортани и к 7-й неделе достигает места окончательной локализации, формируя две доли и перешеек.
- Ductus thyroglossus. Зачаток железы сначала связан с глоткой при помощи полого тяжа, открывающегося на поверхности корня языка (позднее —foramen соесит). Нормально этот тяж дегенерирует.
(а) Кисты. При неполной дегенерации эпителиального тяжа по его протяжению могут возникать шейные кисты.
(б) Дополнительные желёзки. Часть клеток тяжа может образовать функционально активные островки эндокринной ткани щитовидной железы.
(в) Пирамидальная доля железы — наиболее близкий к телу железы остаток тяжа.
- Специфическая дифференцировка. Транскрипционный фактор TTFl (белок с Mr 35 кД) активирует гены тироглобулина, тиропероксидазы, рецепторов ТТГ и jV-ацетилглюкозамина.
- Пренатальная железа. В конце третьего месяца развития плода начинается синтез йодсодержащих гормонов, появляющихся в амниотической жидкости.
(а) Содержание Tj + T1 амниотической жидкости меньше T3 + T4 крови матери.
(б) Содержание г T3 (сМ. IV В I в (6)) в амниотической жидкости много выше такового в крови матери. Это обстоятельство означает, что преобладающий йодсодержащий гормон плода — гТ3.
Измерение гT3 в амниотической жидкости используют для диагностики возможной недостаточности функции щитовидной железы (гипотиреоидизм плода).
в. Вилочковая железа. Третья и четвёртая пары глоточных карманов.
- Нервный гребень. Синтезирующие кальцитонин светлые (С-клетки) щитовидной железы развиваются из клеток нервного гребня.
Б. Строение (рис. 9-13). Как и всякий орган, щитовидная железа складывается из двух основных частей — стромы и паренхимы.
- Строма состоит из вспомогательных структур.
а. Капсула сформирована из плотной волокнистой соединительной ткани.
б. От капсулы отходят тяжи (стандартное наименование — септы, или трабекулы) плотной волокнистой соединительной ткани, содержащие кровеносные и лимфатические сосуды, нервы.
в. Интерстиций. Пространство органа заполняет поддерживающий элементы паренхимы каркас из рыхлой волокнистой соединительной ткани с кровеносными и лимфатическими сосудами, отдельными нервными волокнами и их окончаниями.
- Кровоток железы интенсивен и сопоставим с кровоснабжением мозга, перфузией крови через почки и печень.
- Кровеносные капилляры фенестрированного типа контактируют с эндокринными клетками паренхимы.
- Иннервация
(а) Соматическая чувствительная. В железе найдены чувствительные нервные окончания, образованные ветвлениями периферических отростков чувствительных нейронов.
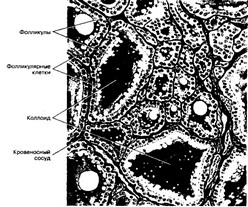
Рис. 9-13. Щитовидная железа [из Stdhr Р, Mdllendoiff Ws 1933]
(б) Двигательная вегетативная (симпатическая и парасимпатическая). Преобладают сопровождающие кровеносные сосуды и иннервирующие их ГМК варикозные ветвления постганглионарных симпатических нейронов. Влияние вегетативной иннервации на эндокринную функцию незначительно.
- Паренхима — совокупность гистологических элементов, выполняющих основную функцию органа. Паренхима щитовидной железы (эндокринная функция) — совокупность секретирующих тиреоидные гормоны клеток и С-клеток, синтезирующих кальцитонин. И те, и другие входят в состав фолликулов и скоплений межфолликулярных клеток. ¦
а. Фолликулы — различной величины и формы (преимущественно округлые) образования, содержащие комоид. Стенка фолликула образована эпителиальными фолликулярными клетками (синтез T4 и T3), прикреплёнными к базальной мембране. Между базальной мембраной и фолликулярными клетками встречаются более крупные светлые клетки (синтез кальцитонина).
- Фолликулярные клетки (тироциты, рис. 9-14) образуют стенку фолликула и формируют его содержимое, синтезируя и секретируя в коллоид тироглобулин. Тиропероксидаза и рецептор jV-ацетилглюкозамина (другие специфические для фолликулярных клеток белки) также синтезируются в фолликулярных клетках,
(а) Функции. Основная функция фолликулярных клеток — синтез и секреция T4 и T3 — складывается из многих процессов:
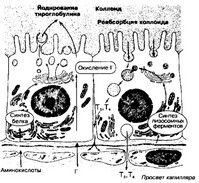
Рис. 9-14. Участие фолликулярных клеток в секреции йодсодержащих гормонов [из Junqueira LC, CameiroJ, 1991]
- поглощения йода,
- окисления йода,
- образования тироглобулина,
- секреции тироглобулина в полость фолликула,
- йодирования тироглобулина,
- эндоцитоза и расщепление тироглобулина,
- секреции T3 и T4.
(б) Морфология
- Форма клеток (от низкокубической до цилиндрической) эпителиальной стенки фолликула зависит от интенсивности их функционирования: высота клеток пропорциональна напряжённости осуществляемых в них процессов. Тиротропин стимулирует функцию фолликулярных клеток.
- Полярная дифференцировка эпителиальных клеток выражена хорошо.
- Базальная часть клеток содержит ядро, гладкий и шероховатый эндо- плазматический ретикулум. В плазмолемму вмонтированы рецепторы ТТГ. Возможна складчатость плазмолеммы (отражает интенсивность обмена между клетками и кровеносными капиллярами — захват йода, поступление метаболитов, секреция гормонов).
- Латеральная часть клеток содержит межклеточные контакты, предупреждающие просачивание коллоида.
- Апикальная часть содержит выраженный комплекс Гольджи (формирование секреторных пузырьков, присоединение углеводов к тироглобулину), разные
типы пузырьков (секреторные [содержат тироглобулин), окаймлённые [незрелый тироглобулнн из полости фолликула поступает в клетку для йодирования], эндоцитозные [содержат зрелый тироглобулин для его последующей деградации в фаголизосомах[), микроворсинки (увеличение поверхности обмена между клетками и полостью фолликула). Апикальная плазмолемма содержит рецепторы TV-ацетилглюкозамина (связывание незрелого тироглобулина для его интернализации путём опосредованного этими рецепторами эндоцитоза). В связи с мембранными структурами апикальной части клеток находится тиропероксидаза.
- Митохондрии, лизосомы, фаголизосомы рассеяны по цитоплазме.
- Термин коллоид точно определяет физико-химическую природу содержимого фолликулов. Главный компонент — тироглобулин различной степени зрелости.
(а) Незрелый тироглобулин (нейодированный и частично йодированный) секрети- руется тироцитами в полость фолликула.
(б) Созревание тироглобулина происходит примерно в течение двух суток в апикальной части фолликулярных клеток путём его йодирования при помощи тиропероксидазы.
(в) Термин рециклизация тироглобулина обозначает циклы интернализации незрелого тироглобулина из коллоида в апикальную часть тироцитов, йодирование тироглобулина и его секрецию в коллоид.
Интернализация тироглобулина происходит путём опосредованного рецепторами jY-ацетилглюкозамина эндоцитоза.
(г) Зрелый тироглобулин (полностью йодированный) — прогормон йодсодержащих гормонов, форма их хранения в коллоиде. По мере необходимости зрелый тироглобулин фагоцитируется тироцитами, поступает в лизосомы и расщепляется в фаголизосомах. Аминокислоты используются для новых синтезов, a T3 и T4 из базальной части клеток поступают в кровь.
- С-клетки (произносят «си-клетки», от англ. calcitonin, кальцитонин) в составе фолликулов называют также парафолликулярными клетками. В них происходит экспрессия кальцитонинового гена CALC1, кодирующего кальцитонин, катакаль- цин и относящийся к кальцитониновому гену пептид а. С-клетки крупнее тироцитов, в составе фолликулов расположены, как правило, одиночно. Морфология этих клеток характерна для клеток, синтезирующих белок на экспорт (присутствуют шероховатая эндоплазматическая сеть, комплекс Гольджи, секреторные гранулы, митохондрии). На гистологических препаратах цитоплазма С-клеток выглядит светлее цитоплазмы тироцитов, отсюда их название — светлые клетки.
- Хюртля клетки. Иногда (например, при болезни Хашимото) в составе стенки фолликулов или между фолликулами находят крупные клетки с зернистой окси- фильной цитоплазмой, содержащие много митохондрий — онкоциты, или клетки Хюртля (Гюртля, также Асканази-Хюртля).
К сожалению, по отношению к этим клеткам наблюдается терминологическая путаница. Иногда их именуют парафолликулярные (т.е. С-клетки); более того, в части источников клетками Асканази-Хюртля называют продуцирующие кальцитонин клетки,
б. Межфолликулярные клетки. К паренхиме щитовидной железы, помимо образующих фолликулы клеток, относятся также островки клеток, расположенные между фолликулами. Островки образованы способными синтезировать йодсодержащие гормоны клетками (малодифференцированные тироциты, формирующие новые фолликулы), а также С-клетками.
В. Гормоны. Щитовидная железа секретирует йодсодержащие гормоны и продукты экспрессии кальцитонинового гена (в т.ч. кальцитонин).
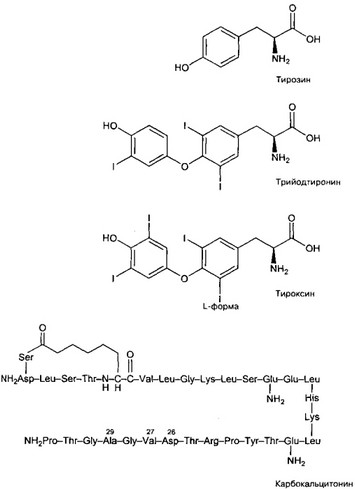
- Йодсодержащие гормоны — T4 и T3 (рис. 9-15) — образуются в составе тироглобулина при йодировании тирозила (формируются монойодтирозил и дийодтирозил). Затем ти- роглобулин расщепляется в фаголизосомах до реутилизируемых клеткой аминокислот, а из монойодтирозина и дийодтирозина образуются трийодтиронин [T3] и тетрайодтиронин [T4]. Этот процесс, а также йодирование тирозина катализирует тиропероксидаза. Далее йодированные соединения выделяются из клетки.
а. Монойодтирозин образуется при введении йода по одной позиции тирозила.
б. Дийодтирозин образуется при введении йода по двум позициям тирозила. Гормональной активностью, как и монойодтирозин, не обладает; оба соединения выделяются из фолликулярных клеток, но быстро захватываются обратно и дейодинируются.
в. Тироксин {р-[(3,5-дийодо-4-гидроксифенокси)-3,5-дийодофенил]аланин, или 3,5,3',5'- тетрайодтиронин, C15H11I4NO4, мол. масса 776,87} образуется из пары дийодгирози- нов. Тироксин — основной йодсодержащий гормон, на долю T4 приходится не менее 90% всего содержащегося в крови йода.
- Транспорт в крови. He более 0,05% T4 циркулирует в крови в свободной форме, практически весь тироксин находится в связанной с белками плазмы форме. Главный транспортный белок — тироксин-связывающий глобулин (связывает 80% Т.), на долю тироксин-связывающего преальбумина, а также альбумина приходится 20% T4.
- Время циркуляции в крови (время полужизни) T4 около 7 дней, при гиперти- реоидизме — 3-4 дня, при гипотиреоидизме — до 10 дней.
- L-форма тироксина физиологически примерно вдвое активнее рацемической (DL-тироксин), D-форма гормональной активности не имеет.
- Дейодирование наружного кольца тироксина (рис. 9-15), частично происходящее в щитовидной железе, осуществляется преимущественно в печени и приводит к образованию T3.
- Дейодирование внутреннего кольца тироксина (рис. 9-15) происходит в щитовидной железе, преимущественно в печени и частично в почке, в результате образуется т.н. реверсивный (обратный) T3 (3,3',5’-трийодтиронин, гТ3 [от англ. reverse]), имеющий незначительную физиологическую активность.
г. Трийодотиронин [3,5,3'-трийодтиронин, или 4-(3-йодо-4-гидроксифенокси)-3,5-дий- одофенилаланин, C15H12I3NO4, мол. масса 650,98] образуется из монойодтиронина и дийодтиронина. На долю T3 приходится лишь 5% содержащегося в крови йода, но T3 не менее важен, чем тироксин, для реализации эффектов йодсодержащих гормонов.
- Транспорт в крови. He более 0,5% T3 циркулирует в крови в свободной форме, практически весь трийодтиронин находится в связанной форме.
- Физиологическая активность T3 примерно в четыре раза выше, чем тироксина, но время полужизни вдвое меньше.
- В щитовидной железе образуется около 15% циркулирующего в крови T3. Остальной трийодтиронин образуется при монодейодировании наружного кольца тироксина, происходящем преимущественно в печени.
д. Функции йодсодержащих гормонов многочисленны. Например, T3 и T4 увеличивают обменные процессы, ускоряют катаболизм белков, жиров и углеводов, необходимы для нормального развития ЦНС, увеличивают частоту сердечных сокращений и сердечный выброс. Крайне разнообразные эффекты йодсодержащих гормонов на клетки- мишени (ими практически являются все клетки организма) объясняют увеличением синтеза белков и потребления кислорода.
- Синтез белков увеличивается в результате активации транскрипции в клетках- мишенях.
- Потребление кислорода возрастает в результате увеличенной активности Na+,K+- АТФазы.
е. Рецепторы тиреоидных гормонов относят к факторам транскрипции. Известно не менее трёх подтипов: а,, а2, р. Подтипы а, и р — трансформирующие гены ERBAl и ERBA2 соответственно.
- Подтип а, (онкоген ERBA1, 190120, 17q 11.2) экспрессируется преимущественно в ЦНС, практически отсутствует в печени.
Кретинизм. Имеется аутосомно-доминантная форма врождённого кретинизма.
- Подтип а2 экспрессируется в печени, почке, плаценте, мозге и других органах. Врождённая глухота. Известны мутации гена, приводящие, в частности, к глухоте и дефектам скелета.
- Подтип р (онкоген ERBA2, 190160, 3р24.3) экспрессируется во многих органах.
Нечувствительность к тиреоидным гормонам. Известно около 40 дефектов гена, приводящих на фоне различной выраженности гипертиреоидизма к развитию различных синдромов.
- Гиперактивности синдром
- Периодическая тахикардия
- Затруднённое обучение
- Низкорослость
- Глухота
ж. Оценка функций щитовидной железы должна проводиться по многим параметрам.
- Радиоиммунологический анализ позволяет прямо измерять содержание T3, T4, ТТГ. При
этом следует учитывать соотношение между свободными и связанными формами гормонов.
- Поглощение гормонов смолами — широко используемый непрямой метод определения связывающих гормоны белков.
- Индекс свободного тироксина — оценка свободного T4C учётом содержания связывающих гормоны белков.
- Тест стимуляции ТТГ тиролиберином определяет секрецию в кровь тиротропина в ответ на внутривенное введение тиролиберина.
- Тесты выявления AT к рецепторам ТТГ выявляют гетерогенную группу Ig, связывающихся с рецепторами ТТГ эндокринных клеток щитовидной железы и изменяющих её функциональную активность.
Аутоантитела обнаружены и к другим белкам щитовидной железы (например, к тиропероксидазе). Имеются также наследуемые формы аутоиммунных заболеваний щитовидной железы.
- Сканирование щитовидной железы при помощи изотопов технеция (99mTc) позволяет выявить области пониженного накопления радионуклида (холодные узлы), обнаружить эктопические очаги щитовидной железы или дефект паренхимы органа. 99nTc накапливается только в щитовидной железе, период полувыведения составляет всего 6 часов.
- Исследование поглощения радиоактивного йода при помощи йода-123 (123I) и йода-131 (131I).
- Содержание йода в питьевой воде. Проводится йодирование воды на водопроводных станциях.
з. Тиреоидный статус определяет эндокринную функцию щитовидной железы. Эутирео- идия — отсутствие отклонений. Заболевание щитовидной железы можно предположить при появлении симптомов недостаточности эндокринной функции (гипотиреоз), избыточных эффектов тиреоидных гормонов (гипертиреоз) либо при очаговом или диффузном увеличении щитовидной железы (зоб).
- Гипотиреоз
(а) Врождённый (первичный) гипотиреоидизм — относительно частая патология (I на 4000 новорождённых). Раннее выявление патологии может предотвратить развитие серьёзных неврологических осложнений. Разработаны методы массового обследования тиреоидного статуса. В отечественной практике оценка проводится по уровню ТТГ на пятый день жизни.
(б) Ювенильный (приобретённый) гипотиреоидизм. Приобретённым называют гипо- тиреоидизм, проявляющийся после первого года жизни. Ювенильный гипотиреоидизм чаще встречается у девочек. Наиболее распространённая причина — аутоиммунное поражение щитовидной железы. Для лечения проводят заместительную терапию (синтетический Х-тироксин).
(в) Хронический тиреоидит — наиболее частая причина гипотиреоза у взрослых.
- Гипертиреоз
(а) Болезнь Грейвса (диффузный токсический зоб) — наиболее частая причина гиперти- реоза. Это аутоиммунное заболевание, при котором тиреоид-стимулирующие Ig связываются с рецепторами ТТГ на фолликулярных клетках щитовидной железы, что приводит к диффузному увеличению железы и стимуляции выработки тиреоидных гормонов.
(б) Болезнь Пламмера (узловой токсический зоб) встречается реже, чем болезнь Грейвса, и обычно у пожилых лиц.
- Зоб
Тиреоидит Хасимото (хронический лимфоцитарный тиреоидит) — частое аутоиммунное заболевание, поражающее в основном женщин. У большинства больных находят антитиреоидные AT.
- Узелки в щитовидной железе выявляют у 1% лиц в возрасте около 20 лет и у 5% лиц в возрасте около 60; рак обнаруживают в 10-20% обследованных узелков. Узелки щитовидной железы могут быть аденомами, кистами, локализованными участками хронического тиреоиди- та, коллоидными узелками, геморрагической некротической тканью или карциномой.
- Кальцитонин и другие пептиды кальцитониновых генов CALC.
а. Гены. Известно три гена кальцитонина. CALCl и CALC2 кодируют последовательности Са2+-регулирующих гормонов и относящихся к кальцитониновому гену пептидов. Транскрипты подвергаются альтернативному сплайсингу, что приводит к органоспецифичному синтезу разных пептидов. CALC3— нетранскрибируемый псевдоген.
- Ген CALCl (114130, 11р15.2-р15.1) содержит последовательности пептидных гормонов кальцитонина (экзон 4), катакальцина и (относящегося к кальцитониновому гену) пептида а (экзон 5). В нормальной щитовидной железе экспрессируются последовательности регуляторов Ca2+ — кальцитонина и катакальцина; пептид а нормально в щитовидной железе не экспрессируется.
Медуллярная карцинома щитовидной железы развивается из С-клеток, в её клетках синтезируются все три пептида.
- Ген CALC2 (114160, I lpter-1 Iql2) содержит последовательности разных пептидов, включая кальцитонин и (относящийся к кальцитониновому гену) пептид р. Похоже, CALC2 не транскрибируется в С-клетках.
б. Гормоны
- Кальцитонин — пептид, содержащий 32 аминокислотных остатка, мол. масса 3421. В клинике применяют синтетические аналоги гормона человека, свиньи, лосося.
(а) Карбокальцитонин (рис. 9-15) — синтетический аналог кальцитонина, Cm8H244N12O47, мол. масса 3363,821, значительно устойчивее кальцитонина.
(б) Регулятор экспрессии — Ca2+ плазмы крови, внутривенное его введение существенно увеличивает секрецию кальцитонина.
(в) Функции кальцитонина, как одного из регуляторов кальциевого обмена, определяют как антагонистические функциям гормона паращитовидной железы.
- Уменьшение содержания Ca2* в крови (паратиреокрин увеличивает содержание Ca2+).
- Стимуляция минерализации кости (ПТГ усиливает резорбцию кости),
- Усиление почечной экскреции Ca2+, фосфатов и Na+ (уменьшается их
реабсорбция в канальцах почки).
- Желудочная и панкреатическая секреция. Кальцитонин уменьшает кислотность желудочного сока и содержание амилазы и трипсина в соке поджелудочной железы.
- Гормональная регуляция костной тканя {см. главу 6.3 Б 11).
(г) Рецептор кальцитонина (ген CALCR, 114131, 7q22) относится к семейству рецепторов секретина, при связывании кальцитонина с рецептором в клетках- мишенях (например, остеокласты) происходит увеличение содержания цАМФ.
(д) Семейный полиэндокринный аденоматоз (СПЭА). При некоторых формах СПЭА развивается продуцирующая кальцитонин медуллярная карцинома щитовидной железы.
- Катакальцин — пептид, состоящий из 21 аминокислотного остатка, — имеет те же функции, что и кальцитонин.
- Относящиеся к кальцитониновому гену пептиды а и р (37 аминокислот) экспрессируются в ряде нейронов ЦНС и на периферии (особенно в связи с кровеносными сосудами). Их функции — участие в ноцицепции, пищевом поведении, а также в регуляции тонуса сосудов. Рецепторы к этим пептидам найдены в ЦНС, сердце, плаценте.