
Основную группу клеток-продуцентов цитокинов в адаптивном иммунном ответе представляют лимфоциты. Покоящиеся клетки не секретируют цитокины. При распознавании антигена и при участии рецепторных взаимодействий (CD28-CD80/86 для Т-лимфоцитов и CD40-CD40L для В-лимфоцитов) происходит активация клеток, приводящая к транскрипции генов цитокинов, трансляции и секреции гликозилированных пептидов в межклеточное пространство.
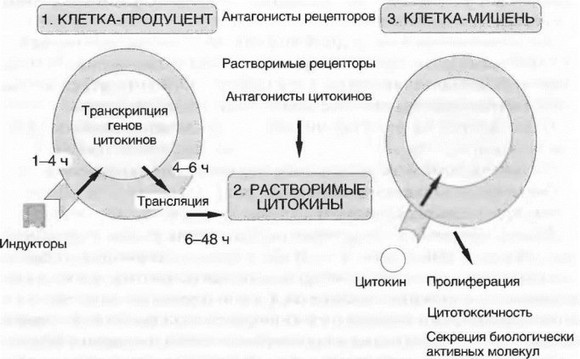
Рис. 7.1. Система цитокинов
CD4 Т-хелперы представлены субпопуляциями: ThO, Thl, Th2, ТЫ7, Tfh, которые различаются между собой спектром секретируе- мых цитокинов в ответ на различные антигены.
ThO вырабатывают широкий спектр цитокинов в очень низких концентрациях.
Направление дифференцировки ThO определяет развитие двух форм иммунного ответа с преобладанием гуморальных или клеточных механизмов.
Природа антигена, его концентрация, локализация в клетке, тип антигенпрезентирующих клеток и определенный набор цитокинов регулируют направление дифференцировки ThO.
Дендритные клетки после захвата и процессинга антигена представляют антигенные пептиды ThO клеткам и вырабатывают цитокины, регулирующие направление их дифференцировки в эффек- торные клетки. Роль индивидуальных цитокинов в данном процессе отражена на рис. 7.2. ИЛ-12 индуцирует синтез ИФНу Т-лимфоцитами и NK. ИФНу обеспечивает дифференцировку ТЫ, которые начинают секретировать цитокины (ИЛ-2, ИФНу, ИЛ-3, ФНОа, лимфотокси- ны), регулирующие развитие реакций на внутриклеточные патогены
(гиперчувствительности замедленного типа (ГЗТ) и различные типы клеточной цитотоксичности).
ИЛ-4 обеспечивает дифференцировку ТЪО в Th2. Активированные Th2 вырабатывают цитокины (ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-13 и др.), определяющие пролиферацию В-лимфоцитов, их дальнейшую дифференцировку в плазматические клетки,и развитие реакций анти- телогенеза, преимущественно на внеклеточные патогены.
ИФНу негативно регулирует функцию ТЪ2-клеток и, наоборот, ИЛ-4, ИЛ-10, секретируемые Th2, угнетают функцию Thl (рис. 7.3). Молекулярный механизм этой регуляции связан с транскрипционными факторами. Экспрессия T-bet и STAT4, детерминированная ИФНу, направляет дифференцировку Т-клеток по пути ТЫ и супрессирует развитие Th2. ИЛ-4 индуцирует экспрессию GATA-3 и STAT6, что соответственно обеспечивает превращение наивных ThO в Тй2-клетки (рис. 7.2).
В последние годы описана особая субпопуляция Т-клеток хелпе- ров (ТЫ7), продуцирующих ИЛ-17. Члены семейства ИЛ-17 могут экспрессироваться активированными клетками памяти (CD4CD45RO), у5Т-клетками, NKT клетками, нейтрофилами, моноцитами под влиянием ИЛ-23, ИЛ-6, ТФРр, вырабатываемых макрофагами и дендритными клетками. Основным дифференцировочным фактором у человека является ROR-C, у мышей — ROR-yi. Показана кардинальная роль ИЛ-17 в развитии хронического воспаления и аутоиммунной патологии (см. рис. 7.2).
Кроме того, Т-лимфоциты в тимусе могут дифференцироваться в естественные клетки-регуляторы (Treg), экспрессирующие поверхностные маркеры CD4+ CD25+ и транскрипционный фактор FOXP3. Эти клетки способны подавлять иммунный ответ, опосредуемый ТЫ и Т112-клетками, путем прямого межклеточного контакта и синтеза ТФРР и ИЛ-10.
Схемы дифференцировки клонов ThO и секретируемых ими цитокинов представлены на рис. 7.2 и 7.3 (см. также цв. вклейку).
Т-цитотоксические клетки (CD8+), естественные киллеры — слабые продуценты цитокинов, таких, как интерфероны, ФНОа и лим- фотоксины.
Избыточная активация одной из субпопуляций Th может определить развитие одного из вариантов иммунного ответа. Хроническая несбалансированность активации Th способна привести к формированию иммунопатологических состояний, связанных с проявления-
ми аллергии, аутоиммунной патологии, хронических воспалительных процессов и др.
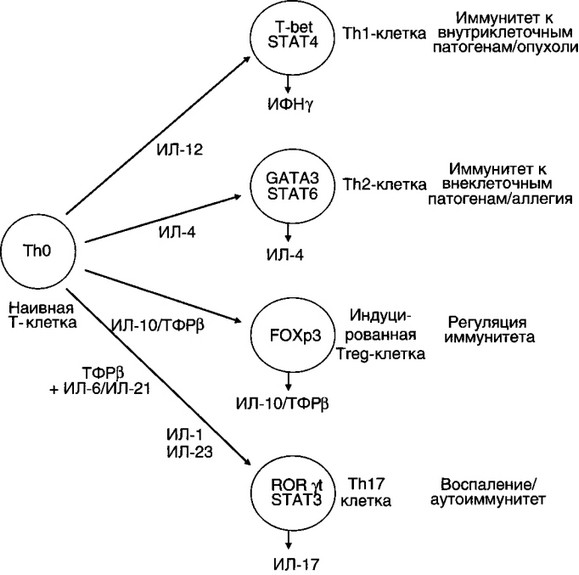
Рис. 7.2. Различные субпопуляции Т-лимфоцитов, продуцирующие цитокины
II. В системе врожденного иммунитета основными продуцентами цитокинов являются клетки миелоидного ряда. С помощью ТоИ-по- добных рецепторов (TLRs) они распознают сходные молекулярные структуры различных патогенов, так называемые патогенассоцииро- ванные молекулярные патерны (РАМП), например липополисахарид (ЛПС) грамотрицательных бактерий, липотейхоевые кислоты, пепти- догликаны грамположительных микроорганизмов, флагеллин, ДНК, богатую неметилированными CpG повторами, и др. В результате
такого взаимодействия с TLR запускается внутриклеточный каскад передачи сигнала, приводящий к экспрессии генов двух основных групп цитокинов: провоспалительных и ИФН типа 1 (рис. 7.4, см. также цв. вклейку). Главным образом эти цитокины (ИЛ-1, -6, -8, -12, ФНОа, ГМ-КСФ, ИФН, хемокины и др.) индуцируют развитие воспаления и участвуют в защите организма от бактериальных и вирусных инфекций.
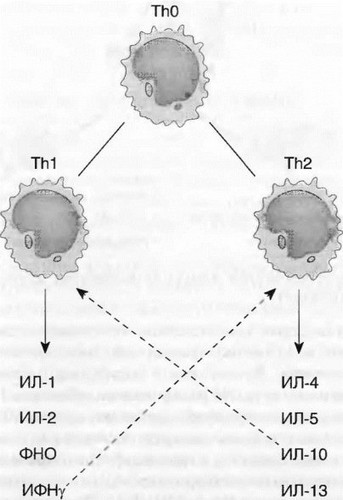
Рис. 7.3. Спектр цитокинов, секретируемых ТЫ- и ТИ2-клетками
Цитокины и их антагонисты подробно описаны в ряде монографий (Ковальчук Л.В. и соавт., 2000; Кетлинский С.А., Симбирцев А.С., 2008).
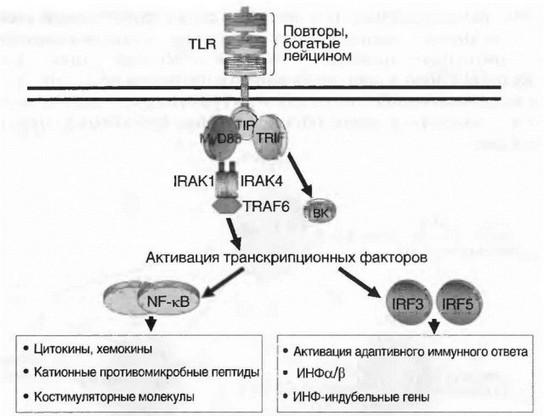
Рис. 7.4. TLR-опосредованная индукция выработки цитокинов клетками врожденного иммунитета
Избыточная экспрессия цитокинов небезопасна для организма и может привести к развитию чрезмерной воспалительной реакции, острофазового ответа. В регуляции выработки провоспалительных цитокинов принимают участие различные ингибиторы. Так, описан ряд веществ, которые неспецифически связывают цитокин ИЛ-1 и препятствуют проявлению его биологического действия (а2-макроглобулин, СЗ-компонент комплемента, уромодулин). Специфическими ингибиторами ИЛ-1 могут быть растворимые рецепторы-ловушки, антитела и рецепторный антагонист ИЛ-1 (ИЛ-IRA). При развитии воспаления происходит усиление экспрессии гена ИЛ-IRA. Но и в норме этот антагонист присутствует в крови в высокой концентрации (до 1 нг/мл и более), блокируя действие эндогенного ИЛ-1.
Рис. 7.1. Система цитокинов
CD4 Т-хелперы представлены субпопуляциями: ThO, Thl, Th2, ТЫ7, Tfh, которые различаются между собой спектром секретируе- мых цитокинов в ответ на различные антигены.
ThO вырабатывают широкий спектр цитокинов в очень низких концентрациях.
Направление дифференцировки ThO определяет развитие двух форм иммунного ответа с преобладанием гуморальных или клеточных механизмов.
Природа антигена, его концентрация, локализация в клетке, тип антигенпрезентирующих клеток и определенный набор цитокинов регулируют направление дифференцировки ThO.
Дендритные клетки после захвата и процессинга антигена представляют антигенные пептиды ThO клеткам и вырабатывают цитокины, регулирующие направление их дифференцировки в эффек- торные клетки. Роль индивидуальных цитокинов в данном процессе отражена на рис. 7.2. ИЛ-12 индуцирует синтез ИФНу Т-лимфоцитами и NK. ИФНу обеспечивает дифференцировку ТЫ, которые начинают секретировать цитокины (ИЛ-2, ИФНу, ИЛ-3, ФНОа, лимфотокси- ны), регулирующие развитие реакций на внутриклеточные патогены
(гиперчувствительности замедленного типа (ГЗТ) и различные типы клеточной цитотоксичности).
ИЛ-4 обеспечивает дифференцировку ТЪО в Th2. Активированные Th2 вырабатывают цитокины (ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-13 и др.), определяющие пролиферацию В-лимфоцитов, их дальнейшую дифференцировку в плазматические клетки,и развитие реакций анти- телогенеза, преимущественно на внеклеточные патогены.
ИФНу негативно регулирует функцию ТЪ2-клеток и, наоборот, ИЛ-4, ИЛ-10, секретируемые Th2, угнетают функцию Thl (рис. 7.3). Молекулярный механизм этой регуляции связан с транскрипционными факторами. Экспрессия T-bet и STAT4, детерминированная ИФНу, направляет дифференцировку Т-клеток по пути ТЫ и супрессирует развитие Th2. ИЛ-4 индуцирует экспрессию GATA-3 и STAT6, что соответственно обеспечивает превращение наивных ThO в Тй2-клетки (рис. 7.2).
В последние годы описана особая субпопуляция Т-клеток хелпе- ров (ТЫ7), продуцирующих ИЛ-17. Члены семейства ИЛ-17 могут экспрессироваться активированными клетками памяти (CD4CD45RO), у5Т-клетками, NKT клетками, нейтрофилами, моноцитами под влиянием ИЛ-23, ИЛ-6, ТФРр, вырабатываемых макрофагами и дендритными клетками. Основным дифференцировочным фактором у человека является ROR-C, у мышей — ROR-yi. Показана кардинальная роль ИЛ-17 в развитии хронического воспаления и аутоиммунной патологии (см. рис. 7.2).
Кроме того, Т-лимфоциты в тимусе могут дифференцироваться в естественные клетки-регуляторы (Treg), экспрессирующие поверхностные маркеры CD4+ CD25+ и транскрипционный фактор FOXP3. Эти клетки способны подавлять иммунный ответ, опосредуемый ТЫ и Т112-клетками, путем прямого межклеточного контакта и синтеза ТФРР и ИЛ-10.
Схемы дифференцировки клонов ThO и секретируемых ими цитокинов представлены на рис. 7.2 и 7.3 (см. также цв. вклейку).
Т-цитотоксические клетки (CD8+), естественные киллеры — слабые продуценты цитокинов, таких, как интерфероны, ФНОа и лим- фотоксины.
Избыточная активация одной из субпопуляций Th может определить развитие одного из вариантов иммунного ответа. Хроническая несбалансированность активации Th способна привести к формированию иммунопатологических состояний, связанных с проявления-
ми аллергии, аутоиммунной патологии, хронических воспалительных процессов и др.
Рис. 7.2. Различные субпопуляции Т-лимфоцитов, продуцирующие цитокины
II. В системе врожденного иммунитета основными продуцентами цитокинов являются клетки миелоидного ряда. С помощью ТоИ-по- добных рецепторов (TLRs) они распознают сходные молекулярные структуры различных патогенов, так называемые патогенассоцииро- ванные молекулярные патерны (РАМП), например липополисахарид (ЛПС) грамотрицательных бактерий, липотейхоевые кислоты, пепти- догликаны грамположительных микроорганизмов, флагеллин, ДНК, богатую неметилированными CpG повторами, и др. В результате
такого взаимодействия с TLR запускается внутриклеточный каскад передачи сигнала, приводящий к экспрессии генов двух основных групп цитокинов: провоспалительных и ИФН типа 1 (рис. 7.4, см. также цв. вклейку). Главным образом эти цитокины (ИЛ-1, -6, -8, -12, ФНОа, ГМ-КСФ, ИФН, хемокины и др.) индуцируют развитие воспаления и участвуют в защите организма от бактериальных и вирусных инфекций.
Рис. 7.3. Спектр цитокинов, секретируемых ТЫ- и ТИ2-клетками
- Клетки, не относящиеся к иммунной системе (клетки соединительной ткани, эпителия, эндотелия), конститутивно секретируют аутокринные факторы роста (ФРФ, ЕФР, ТФРр и др.). и цитокины, поддерживающие пролиферацию гемопоэтических клеток.
Цитокины и их антагонисты подробно описаны в ряде монографий (Ковальчук Л.В. и соавт., 2000; Кетлинский С.А., Симбирцев А.С., 2008).
Рис. 7.4. TLR-опосредованная индукция выработки цитокинов клетками врожденного иммунитета
Избыточная экспрессия цитокинов небезопасна для организма и может привести к развитию чрезмерной воспалительной реакции, острофазового ответа. В регуляции выработки провоспалительных цитокинов принимают участие различные ингибиторы. Так, описан ряд веществ, которые неспецифически связывают цитокин ИЛ-1 и препятствуют проявлению его биологического действия (а2-макроглобулин, СЗ-компонент комплемента, уромодулин). Специфическими ингибиторами ИЛ-1 могут быть растворимые рецепторы-ловушки, антитела и рецепторный антагонист ИЛ-1 (ИЛ-IRA). При развитии воспаления происходит усиление экспрессии гена ИЛ-IRA. Но и в норме этот антагонист присутствует в крови в высокой концентрации (до 1 нг/мл и более), блокируя действие эндогенного ИЛ-1.