
Иммуногенность определяет способность антигена вызывать иммунный ответ независимо от его специфичности. Биологической основой для проявления этого свойства антигенов служат те механизмы развития иммунного ответа, которые предполагают участие, помимо лимфоцитов, определяющих специфическую компоненту реакции, некоторых вспомогательных клеток, а также кооперацию различных типов лимфоцитов. Способность чужеродных веществ запускать весь необходимый клеточный ансамбль и составляет основу их иммуногенности. Иммуногенность антигенов зависит не только от свойств молекул, но и от пути и режима их введения в организм, а также дополнительных воздействий (например, использования адъювантов).
Структурно-химические основы иммуногенности
Антигенами могут быть белки и углеводы. Липиды, нуклеиновые кислоты и другие органические вещества (в некоторых случаях — также неорганические, например, некоторые металлы) эффективны лишь в составе комплексных соединений (например, в комплексе с белками), определяя при этом не иммуногенность, а специфичность антигена (т.е. выполняя
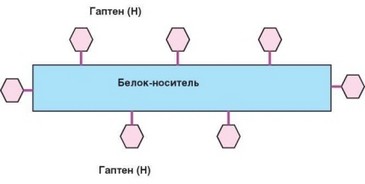
Рис. 3.21. Схема строения комплекса гаптенов с белком-носителем (по К. Ландштей- неру)
роль эпитопа). Использование конъюгатов низкомолекулярных соединений (гаптенов) с белками-носителями (рис. 3.21), введенное в научную практику К. Ландштейнером, сыграло ключевую роль в изучении свойств антигенов и индуцируемых ими антител. В частности, с помощью конъюгатов было показано, что специфичность антигена определяется преимущественно гаптеном, а иммуногенность — носителем. С позиций современной иммунологии, предполагающей необходимость кооперации вовлекаемых в тимусзависимый иммунный ответ В-лимфоцитов с Т-хелперами, иммуно- генность обусловлена преимущественно способностью антигена активировать Т-хелперы. Важным условием развития иммунного ответа служит предварительная активация дендритных клеток в результате распознавания ими PAMP. Именно поэтому следует признать, что иммуногенность молекул антигена во многом определяется наличием в их составе PAMP, т.е. их способностью сформировать при поступлении в организм «провоспалительный фон».
Важнейшее качество, определяющее иммуногенность антигенов, — размер молекулы. С повышением молекулярной массы полимерных молекул увеличивается их иммуногенность. Для полисахаридов это правило срабатывает только в ограниченной степени. Универсальной шкалы зависимости иммуногенности от молекулярной массы не существует. Для углеводов минимальный размер молекулы, обладающей иммуногенностью, больше, чем для белков. Минимальный размер белковой молкулы, вероятно, определяется появлением а-спиральной структуры (7—10 аминокислотных остатков), однако он варьирует в зависимости от конкретного состава молекулы. Минимальная описанная молекулярная масса иммуногенной молекулы составляет 450 Да (арсанил-N-ацетил-DL-тирозин). При переходе от мономерной формы флагеллина (40 кДа) к полимерной (20 000 кДа) титры антител возрастают на 2 порядка. Для углеводов граница между низкой и высокой иммуногенностью расположена на уровне молекулярной массы в десятки килодальтон: полимер декстрана массой 52,3 кДа — слабо иммуногенен, а массой 90,7 кДа вызывает достаточно сильный ответ. Число
антителообразующих клеток в селезенке при иммунизации полимерами пневмококкового полисахарида массой 220, 121 и 40 кДа различается примерно на порядок (со снижением молекулярной массы уровень ответа убывает).
Помимо формирования структур, определяющих иммуногенность (например, а-спирали для белков), размер молекулы важен и для увеличения числа групп (эпитопов), распознаваемых рецепторами лимфоцитов, т.е. для повышения валентности антигена. Значение этого фактора наиболее четко проявляется при использовании конъюгатов, содержащих различное число гаптенных групп. С повышением числа идентичных групп имму- ногенность конъюгата растет, даже если его размеры не увеличиваются. После достижения определенной эпитопной плотности дальнейшее возрастание иммуногенности с увеличением числа эпитопов прекращается и может наблюдаться даже снижение иммуногенности вследствие стеричес- ких помех, создаваемых расположением эпитопов, их взаимной маскировки и блокады факторов, определяющих иммуногенность.
Влияние валентности на иммуногенность связано также с разнообразием эпитопов, присутствующих на молекуле. Установлено, что молекула приобретает иммуногенность только при достаточном разнообразии ее структуры. Так, поли-L-лизин иммуногенен только для ограниченного числа животных, например, для некоторых линий морских свинок. Однако введение в состав этой молекулы боковых цепей или чередование лизина с другими остатками в составе основной цепи делает полимер иммуногенным практически для любых животных.
Наконец, роль размера молекулы в проявлении ее иммуногенности можно проиллюстрировать на примере молекулярных агрегатов. Их высокая иммуногенность в значительной степени обусловлена тем, что они активно фагоцитируются, что важно для процесса обработки антигена и его представления (презентации) Т-хелперам.
Иммуногенность антигенов зависит от жесткости их структуры, т.е. способности сохранять определенную конфигурацию. Стабилизации конформации способствует наличие ароматических заряженных полярных аминокислотных остатков. Так, молекула желатина, утратившая жесткость конформации в результате обработки, практически не иммуногенна, но приобретает иммуногенность после введения в ее состав ароматических аминокислот. Наоборот, гидрофобные остатки в большом количестве препятствуют формированию а-спирали и стабилизации конформации молекул. Чрезмерную гибкость придают полимерам остатки пролина, особенно повторяющиеся. В обоих случаях снижение жесткости молекул сопровождается ослаблением их иммуногенности. Однако при распознавании антигенов достаточно высокая гибкость полимера позволяет эпитопу «подстроиться» под структуру активного центра рецептора В-клеток.
Существует еще одно свойство антигенов, от которого зависит их имму- ногенность: они должны принадлежать к классам полимеров, из которых построены организмы высших животных. Полагают, что это обусловлено необходимостью деградации молекулы антигена для формирования на его основе лиганда для рецепторов Т-клеток (см. раздел 3.2.2.2). Осуществление деградации требует наличия соответствующих ферментов. Ферменты в организме обычно участвуют в расщеплении продуктов питания и собственных макромолекул. Отсутствие ферментов, способных расщеплять некоторые полимеры, служит основой слабой иммуногенности таких веществ. Считают, что с повышением резистентности к расщеплению ферментами связано ослабление иммуногенности белков после их рацемизации щелочами. Таким образом, хотя антигены по определению должны быть чужеродны для организма, эта чужеродность не должна переходить определенные границы. Вышесказанное относится преимущественно к антигенам, вызывающим тимусзависимый ответ.
Генетические аспекты иммуногенности
Поскольку иммуногенность антигена зависит от эффективности процессов обработки, которой он подвергается в организме, следует ожидать, что она зависит и от генотипа реципиента. Существование генетического контроля иммунного ответа на конкретные антигены показано в разнообразных экспериментах. Так, при иммунизации инбредных (генетически «чистых» линий) морских свинок полимером (Glu—Lys)n или конъюгатом динитрофе- нил-поли L-лизином животные одной линии отвечали образованием антител на оба конъюгата, а морские свинки другой линии не отвечали ни на один из них. Гибридологический анализ показал, что способность отвечать на антиген детерминируется одним доминантным геном.
Аналогичные результаты получены в экспериментах на мышах с использованием других синтетических полипептидов. Если мышей иммунизировать разветвленными полипептидами (T, G)—A-L и (His, G)—A-L (эти полипептиды содержат основную полипептидную цепь, образованную поли-Ь-лизином с боковыми полиаланиновыми цепями, заканчивающимися остатками тирозина и глутаминовой кислоты или гистидина и глутаминовой кислоты соответственно), то мыши линии C57BL/6 дают высокий иммунный ответ на 1-й, но не на 2-й пептид. Мыши линии СВА, наоборот, сильно отвечают на 2-й и слабо — на 1-й пептид. И в этом случае сильный ответ детерминируется одним доминантным геном.
И у морских свинок, и у мышей показана связь уровня иммунного ответа с комплексом MHC. Связь иммуногенности с MHC обусловлена различиями в сродстве аллельных вариантов молекул MHC к различным пептидным фрагментам антигенов, выступающих в качестве эпитопов для Т-клеток. Уровень иммунного ответа в значительной степени определяется эффективностью презентации антигенного пептида, а она зависит от способности пептидов встраиваться в молекулы MHC. Таким образом, генетическая детерминация структуры MHC (точнее, участка, связывающего пептид) одновременно распространяется на контроль уровня иммунного ответа.
В такой четкой форме генетическая детерминация иммуногенности проявляется только в отношении некоторых достаточно простых по структуре антигенов. Способность отвечать на сложные антигены подвержена комплексному генетическому контролю (см. раздел 3.6.6.1). Данные генетических исследований подчеркивают относительность понятия «иммуногенность» и зависимость иммуногенности от свойств организма, в который введен антиген. Оказывается, что иммуногенность антигена может зависеть от особенностей реакции организма в не меньшей степени, чем от структуры антигена.
Тимусзависмость антигенов
Гуморальный иммунный ответ на белковые антигены обычно требует участия не только В-, но и Т-клеток и ослабляется в отсутствие тимуса (у генетически бестимусных или тимэктомированных вскоре после рождения животных). При этом В-клетки распознают нативный антиген, тогда как Т-клетки — его фрагменты (эпитопы), встроенные в состав молекул MHC (см. далее). Т-клетки могут выступать в качестве клеток-помощников при иммунном ответе только на те молекулы, чьи фрагменты могут встраиваться в состав молекул MHC (например, на белковые антигены). Ответ на другие антигены (например, полисахаридные) осуществляется В-клетками без участия Т-лимфоцитов. Именно поэтому они могут вызывать ответ у бестимусных nude-мышей. Иногда исключение Т-клеток из ответа на антигены обусловлено другими механизмами. Антигены, способные индуцировать иммунный ответ без участия Т-лимфоцитов, называют тимуснезависимыми, или Т-независимыми (ТН).
ТН-антигены, как правило, крупные молекулы (с молекулярной массой порядка 103 кДа). По химической природе это могут быть полисахариды, ЛПС или белки. Они поливалентны, содержат повторяющиеся эпитопы. Полимеризация и агрегирование тимусзависимых антигенов нередко делает их ТН, а конъюгирование полисахаридов с белками, наоборот, переводит полисахаридные антигены из разряда ТН в тимусзависимые. Многие из ТН-антигенов медленно деградируют и длительно персистируют в организме. Для ответа на эти антигены не требуются их обработка и презентация АПК. По этой причине иммунный ответ на такие антигены не контролируется генами МНС. Однако это не означает, что в иммунном ответе на тимуснеза- висимые антигены участвуют только В-лимфоциты. Вспомогательные клетки (макрофаги, дендритные клетки, Т- и NK-лимфоциты) в той или иной степени вносят вклад в иммунный ответ на эти антигены (см. раздел 3.6.4.2). Например, они могут служить источником цитокинов, способствующих пролиферации В-лимфоцитов, например, BAFF — цитокина семейства TNF, играющего ключевую роль в гомеостазе В-лимфоцитов (см. далее).
Важные сведения были получены при изучении ответа на ТН-антигены мышей с мутацией xid, затрагивающей тирозинкиназу btk и блокирующей дифференцировку В-лимфоцитов до стадии, на которой экспрессируется мембранная молекула Lyb-5. Эти мыши, а также новорожденные особи других линий, у которых не успели созреть Lyb-5+ В-клетки, отвечают не на все ТН-антигены. Антигены, на которые способны отвечать мыши с мутацией xid, обозначают как тимуснезависимые антигены 1 типа (ТН-1; Thymus-independent I), а антигены, ответ на которые у этих мышей отсутствует — как тимуснезависимые антигены II типа (ТН-2). В настоящее время установлено, что у Хid-мышей практически отсутствует одна из субпопуляций В-лимфоцитов — CD5+ B^-клетки и специфический ответ на ТН-2 антигены связывают преимущественно с этими клетками. В качестве примера ТН-1 антигенов можно привести большинство бактериальных ЛПС, поли- флагеллин, полисахарид бордетелл, а также их конъюгаты с гаптенами. Важно отметить, что ТН-1 антигены обладают митогенными свойствами в отношении В-клеток. К ТН-2 антигенам относят полисахаридные антигены (в том числе бактериальные), конъюгаты гаптенов с фиколлом, леваном, некоторые разновидности ЛПС, некоторые синтетические антигены (например, поливинилпирролидон).
Основной изотип антител, специфичных к тимуснезависимым антигенам, — IgM; при этом переключения изотипа обычно не происходит, отсутствует «созревание аффинитета» и практически не формируется иммунологическая память и, как следствие, не развивается вторичный иммунный ответ. Известны, однако, исключения из этих правил (например, переключение на IgA в слизистых оболочках).
ТН-антигены не презентируются Т-хелперам в составе MHC. Именно поэтому Т-хелперы не участвуют в ответе на эти антигены. Активацию В-клеток ТН-антигенами лучше рассмотреть на примере ТН-2 антигенов. Способность активировать B-клетки без помощи Т-лимфоцитов обусловлена особенностями структуры ТН-2 антигенов. Важнейшие их свойства, как уже отмечалось, — наличие большого числа повторяющихся эпитопов и достаточно крупный размер молекулы. При связывании ТН-2 антигена происходит кластеризация BCR на поверхности лимфоцита. Это способствует активации киназ, ассоциированных с BCR, и индукции сигнала, достаточного для преодоления порога активации В-лимфоцитов. Вероятно, определенный вклад в активацию ТН-2 специфических клеток вносит распознавание ими PAMP, входящих в состав большинства ТН-антигенов.
ТН-1 антигены активируют В-лимфоциты несколько иначе. Механизм этой активации в настоящее время до конца не известен. Известно, что ТН-1 антигены способны активировать клетки через митогенные и паттернрас- познающие рецепторы. Однако при низких концентрациях антигена такая активация возможна только при его концентрировании на В-клетках при помощи BCR (поэтому в этих условиях на ТН-1 антигены отвечают только клетки специфичных клонов). Не вполне ясно, поступают ли при этом в клетку активационные сигналы от BCR, связавших антиген. Митогенность ТН-1 антигенов проявляется при повышении их концентрации до уровня, при котором для активации В-клеток дополнительное концентрирование антигена при помощи BCR не требуется. Таким образом, в митогенных концентрациях ТН-1 антигены способны активировать все В-клетки, независимо от специфичности их BCR. В настоящее время лучше всего изучен ответ на ТН-1 антиген — ЛПС, который связывается с LBP и CD14 и в составе этого комплекса активирует клетку через TLR-4. Тем не менее, в ответе В-клеток на ТН-1 антигены остается много неясного.
Считают, что степень участия Т-хелперов или иных вспомогательных клеток (например, естественных киллеров) в ответе на ТН-2 антигены выше, чем при ответе на ТН-1 антигены. Показана роль в развитии ответа на эти антигены ряда цитокинов (IL-3, GM-CSF, IFNy, BAFF).
Толерогенность
Альтернатива индукции иммунного ответа при ответе на антигены — развитие неотвечаемости. Это не просто отсутствие ответа, а активное и избирательное его подавление в отношении данного конкретного антигена. На уровне клетки эту реакцию называют анергией, а на уровне организма — иммунологической толерантностью (см. раздел 4.3). Толерантность индуцируется при введении высоких доз белков и полисахаридов при отсутствии молекулярных агрегатов. Для белков может развиться также низкодозная толерантность.
Одно из важных свойств антигенов, способствующих индукции толерантности, — способность избегать поглощения АПК (например, отсутствие молекулярных агрегатов). Проявлению толерогенности способствует также относительно низкая молекулярная масса и высокая эпитопная плотность. Таким образом, одни и те же молекулы могут выступать в качестве иммуногенов и толерогенов или не оказывать действия на иммунную систему в зависимости от их физического состояния (степени агрегированности). Имеет значение также присутствие в микроокружении цитокинов, регулирующих активность дендритных клеток, обрабатывающих и презентирующих антиген Т-лимфо- цитам: IL-12 и IFNy способствуют проявлению иммуногенности, а IL-10 — толерогенности молекул антигенов. Очень важную роль играет степень экспрессии на поверхности дендритных клеток костимулирующих молекул (что, в свою очередь, зависит от наличия PAMP в молекуле антигена) — анергия развивается при слабой экспрессии этих молекул.