
В иммунологии под специфичностью понимают избирательность взаимодействия индукторов и продуктов иммунных процессов, в частности, антигенов и антител. Выше, при рассмотрении структуры антигенраспозна- ющих рецепторов и антител (см. раздел 3.1.1.2) упоминалось, что их сродство к антигенам и способность взаимодействовать с ними связана с четко ограниченным участком — активным центром или антигенсвязывающим участком.
Антигенные детерминанты
Специфическое взаимодействие антител с молекулой антигена связано с относительно небольшим участком ее поверхности, соответствующим по размеру антигенсвязывающему участку рецепторов и антител. Как уже упоминалось, при изучении иммунного ответа на конъюгаты гаптенов с белками-носителями было установлено, что специфичность конъюгата определяют молекулы гаптена, к которым направлены образующиеся антитела. Связь специфичности антигенов с относительно небольшими участками ее поверхности подтвердилась при анализе специфичности природных молекул белков и полисахаридов, а также при изучении ответа на синтетические полипептиды.
В этих работах было установлено, что практически в любой молекуле антигена есть несколько детерминант, или эпитопов — участков, ответственных за взаимодействие с активными центрами антигенраспознающих молекул. Для антигенов с повторяющейся структурой (например, полисахаридов) характерны повторяющиеся однотипные эпитопы. Белкам свойственны разнообразные по своей структуре эпитопы, к каждому из которых могут вырабатываться специфические антитела. В молекуле устанавливается иерархия эпитопов, когда один из них является доминирующим, что проявляется в преимущественном образовании антител против этого эпитопа (явление иммунодоминантности). После искусственного удаления этого эпитопа свойство иммунодоминантности переходит к другому эпитопу.
Связь специфичности антигенов с относительно небольшими фрагментами их молекул объясняется самой природой этой специфичности. Она служит отражением пространственного соответствия (комплементарности) эпитопа и активного центра рецептора. Чем больше это пространственное соответствие, тем выше сродство эпитопа к активному центру рецептора. Распознавание эпитопа рецептором — пусковой момент иммунного ответа. Антитела, продуцируемые в результате такого ответа, соответствуют по своей специфичности рецептору и, следовательно, специфичны к упомянутому выше эпитопу.
Эпитоп представляет собой выпуклость на поверхности макромолекулярной глобулы, соответствующая по величине и конфигурации полости активного центра антитела. Для того, чтобы эпитоп мог проникнуть в полость, размер эпитопа должен быть несколько меньше размеров полости активного центра. Если протяженность активного центра составляет 6 нм, то длина эпитопа — до 4 нм, а его объем — 2—3 нм3. Эпитоп белковой молекулы может содержать 7—15 аминокислотных остатков (это соответствует массе 0,6—1 кДа), а эпитоп углеводной молекулы — 6 моносахаридных остатков. Наибольшую роль в обеспечении специфичности взаимодействия с активным центром играют концевые остатки в составе эпитопа (на долю концевого сахара приходится 39% энергии взаимодействия с антителом, а на долю 6-го остатка — только 6%). Учитывая постепенное убывание вклада мономеров во взаимодействие с активным центром, можно заключить, что суждения о границах эпитопа и его размерности весьма относительны.
Анализ специфичности антител, образуемых одним организмом, а также моноклональных антител, получаемых при иммунизации одного животного, показывает, что против одного эпитопа образуется широкий спектр разных антител. Это еще раз подчеркивает относительность представлений об эпитопной структуре антигенов и невозможность однозначной локализации эпитопов. Эпитопная структура представится еще более размытой, если учесть различия в индивидуальной реакции на антиген различных представителей одного или разных видов животных. Это отсутствие жесткого соответствия между структурами эпитопов и активных центров становится более понятным, если учесть, что в основе механизма формирования антигенраспознающего репертуара рецепторов лимфоцитов лежат случайные процессы (см. раздел 3.1.3). При вторичном иммунном ответе специфичность образующихся антител и их сродство к антигену становится выше за счет особого отбора, которому подвергаются предшественники антителообразующих клеток в ходе иммунного ответа (см. раздел 3.6.2.2).
Специфичность и разрешающая способность распознавания гаптенов антителами первоначально была изучена с помощью реакции задержки. Ее суть состоит в том, что исследуемый свободный гаптен, взаимодействуя с антителом, не вызывает видимых реакций, но препятствует взаимодействию с ними конъюгатов гаптен—белок, способных вызвать такие реакции (образование осадков — преципитатов или агглютинатов клеток). Степень ослабления этих реакций отражает количество гаптена, вступившего во взаимодействие с антителами. Реакция антител и конъюгата гаптена с белком может быть полностью блокирована идентичным свободным гаптеном, частично — родственным, но не идентичным гаптеном и совсем не блокируется гаптеном, кардинально отличающимся от конъюгированного. Такой анализ позволил установить очень высокую способность антител отличать гаптены различного строения. Выяснилось, что антитела различают оптическую конфигурацию углеводов, замещение атомов водорода на кислотные и азотсодержащие группы, но не на галогены. Четко распознаются позиции замещения — орто-, мета-, пара-, особенно при замещении кислотными группами. Более того, распознается заряд детерминанты, которому соответствует противоположный заряд активного центра.
Линейные и конформационные эпитопы
Белковые молекулы имеют сложную пространственную структуру, при этом гидрофильные остатки экспонированны на поверхности, тогда как гидрофобные скрыты в глубине белковой глобулы. Эпитопы, как правило, соответствуют гидрофильным поверхностным структурам, содержащим циклические остатки, которые придают эпитопу пространственную индивидуальность. При свертывании белковой глобулы могут сближаться остатки, отдаленные друг от друга в линейной последовательности. Это обстоятельство определяет существование двух типов эпитопов — линейных и конформационных. Первые образованы линейной последовательностью аминокислотных остатков, вторые — отдаленными друг от друга остатками, сближенными на поверхности белковой глобулы. Нарушение третичной структуры белка, вызванное разрывом дисульфидных связей или денатурацией, приводит к исчезновению конформационных детерминант при полной сохранности линейных.
Существование конформационных детерминант можно продемонстрировать на примере молекулы лизоцима (рис. 3.22). Нативная молекула этого белка содержит петлю, соответствующую последовательностям 60—83, скрепленную дисульфидной связью. При иммунизации лизоцимом петля выступает как иммунодоминантный эпитоп. Разрыв связи приводит к распрямлению петли, ликвидации эпитопа и нарушению взаимодействия с антителами к нативной молекуле. Нативные и линейные эпитопы могут сосуществовать в одной молекуле, как, например, в молекуле миоглобина, содержащей конформационный эпитоп, захватывающий остатки 34, 53 и 113, а также ряд линейных эпитопов (рис. 3.23).
Взаимоотношение линейных и конформационных эпитопов можно наглядно проиллюстрировать на примере синтетических полипептидов, содержащих остатки Glu, Ala и Tyr. В одном из таких пептидов триада Glu—Ala—Tyr повторяется последовательно и многократно, причем полипептидная цепь имеет а-спиральную конфигурацию. В другом полипептиде к линейному пептиду, образованному остатками Ala, «подшивали» боковые группы Glu—Ala—Tyr (рис. 3.24). К пептидам обоих типов получали антитела. Оказалось, что при реакции антител с пептидами не наблюдалось перекрестных реакций, т.е. пептид 1 не реагировал с антителами к пептиду 2 и наоборот. Реакция полипептидов второго типа со своими антителами блокировалась трипептидом Glu—Ala—Tyr. Реакция полипептидов первого типа со своими антителами блокировалась линейными пептидами, образованными как минимум 17 чередующимися остатками Glu, Ala, Tyr, формирующими а-спираль. Таким образом, антитела к линейному полипептиду были
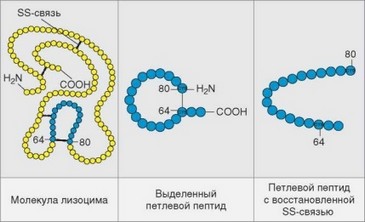
Рис. 3.22. Роль трехмерной структуры молекулы в формировании эпитопа. Пептид 64—80 иммунодоминантный. Он сохраняет способность взаимодействовать с антителами после выделения из молекулы, но утрачивает ее после восстановления дисульфидной связи
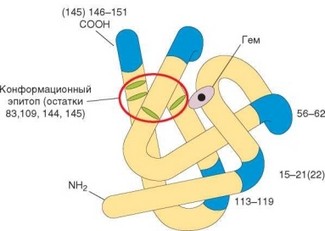
Рис. 3.23. Линейные и конформационные эпитопы (на примере молекулы миоглоби- на кашалота). Синим отмечены линейные эпитопы; красным обведен конформаци- онный эпитоп
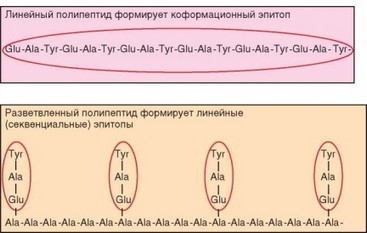
Рис. 3.24. Линейные и конформационные эпитопы модельных синтетических пептидов. Эпитопы обведены красным
направлены к конформационной детерминанте, формируемой а-спиралью, а антитела к разветвленному полипептиду — к линейной детерминанте.
Существование конформационных эпитопов демонстрирует важность распознавания не только линейной структуры, но и пространственной конфигурации молекул. Это положение можно проиллюстрировать на примере перекрестной реактивности гаптенов различной химической природы с белковыми эпитопами антиидиотипических антител (т.е. анти-антител, распознающих некоторые идиотопы антител, специфичных к этим гап- тенам). Перекрестная реактивность в этом случае основана на том, что конфигурация активного центра (и в определенной степени идиотопов) комплементарна конфигурации как эпитопа иммуногена, так и антиидио- топа. Следовательно, эпитоп и антиидиотоп, если не идентичны, то сходны друг с другом по конфигурации.
Анализ специфичности конформационных детерминант с помощью методов, разработанных при изучении гаптенов и применяемых для линейных (секвенциальных) детерминант, практически невозможен. При извлечении из целой молекулы конформационная детерминанта изменяет свою конфигурацию и теряет способность реагировать с антителами. Эти эпитопы изучают, подбирая белки (от родственных видов, мутантных линий клеток и особенно часто — синтетические белки), отличающиеся единичными заменами аминокислотных остатков. При этом для оценки реакции часто используют, твердофазный иммуноферментный анализ, позволяющий достичь высокой производительности и степени стандартизации.
Размеры конформационных эпитопов варьируют даже в более широких пределах, чем линейных эпитопов: они соответствуют 6—17 аминокислотным остаткам. Так, в молекулах миоглобина размер конформационного
эпитопа составляет 6—8, а в молекуле бычьего сывороточного альбумина — 10—12 остатков. Расположение частей конформационных детерминант на большом расстоянии в линейной структуре молекулы описано, например, для аллотипических детерминант иммуноглобулинов, уже упоминавшихся эпитопов миоглобина и т.д. Как в линейных, так и в конформационных эпитопах роль отдельных остатков может существенно варьировать. Так, в формировании описанной выше петлевой детерминанты лизоцима ключевую роль играет остаток в положении 68. В данном случае проявляется фактор гибкости эпитопа, позволяющий «подогнать» конфигурацию кон- формационного эпитопа, к структуре активного центра антител. Подгонка происходит за счет остатков, обеспечивающих эту гибкость (например, пролина).
Важным и практически неизученным остается вопрос об активных факторах, определяющих иерархию эпитопов и реализуемых через ингибирование одними эпитопами иммунного ответа на другие эпитопы, что может происходить с участием регуляторных клеток.
Несмотря на то, что знания о структурных основах конформационных и линейных эпитопов белков пока далеко не полны, их достаточно, чтобы с высокой долей уверенности прогнозировать, какие участки белковой молекулы окажутся антигенными эпитопами. С помощью биоинформатики с использованием компьютерных программ разработаны алгоритмы для таких расчетов. В первую очередь выбирают участки с преобладанием гидрофильных остатков над гидрофобными (условие локализации эпитопа на поверхности молекулы), а также с аминокислотными остатками, придающими этому участку гибкость. Детерминанты, смоделированные на основе таких расчетов, в настоящее время синтезируют и с успехом применяют в серодиагностике и для приготовления искусственных вакцин.