ГЛАВА 3 СИСТЕМА HLA И ИММУНОЛОГИЧЕСКАЯ РЕАКТИВНОСТЬ
Иммуногенетику можно считать самостоятельной наукой с момента открытия комплекса тканевой гистосовместимости у мышей (Н-2). Понятие о главном комплексе гистосовместимости МНС (Major Histo compatibility Complex) сформировалось на основе открытия аналогичных ей систем у ряда других животных. У человека МНС получил название HLA (Human Leukocyti Antigens).
Учение о системе генов, контролирующих синтез антигенов гистосовместимости, за последние десятилетия вышло за рамки трансппан- тационной иммунологии. Многие исследователи ставят вопрос об ответственности МНС за взаимодействие не только между иммуно- компетентными клетками, но и между любыми клетками организма, и о том, обеспечивается ли при этом функциональная целостность последнего. С этих позиций кооперация клеток при иммунологических реакциях представляет собой всего лишь частный случай межклеточ*- ных взаимодействий. Установлено, что антигены МНС человека участвуют в физиологических феноменах, не имеющих прямого отношения к иммунитету: в продукции гормонов, миграции клеток, процессах старения. Антигены МНС экспрессированы не только на иммуноком- петентных, но и на многих других клетках. Гены системы HLA и их антигены принимают самое непосредственное участие на всех стадиях иммунного ответа.
В частности, установлено, что сам факт реакции со стороны иммунной системы на антиген не возможен без системы МНС, ибо как чужеродная распознается не сама молекула антигена, а ее комплекс с собственной молекулой белка (антигеном HLA), являющейся продуктом МНС организма, в который проник антиген. Кооперация клеток в иммунном ответе, участие Т-хелперов и Т-супрессоров обеспечиваются антигенами системы HLA. Прослежена также связь последней с функцией отдельных механизмов иммунной системы. Например, выявлена связь между экспрессией антигенов HLA—А1, В8, DR3 в составе фенотипа и повышенной иммунологической реактивности. Наконец, эффекторные механизмы, в частности Т-киллеры, атакуют мишень при непосредственном участии HLA-антигенов.
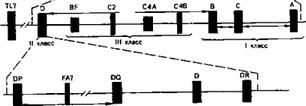
В настоящее время в основном закончено картирование генов системы HLA, которая локализуется на коротком плече справа от
Схема 6. Строение системы HLA [Медуницин Н. В., Алексеев А. Л., 1987]
PGM
V "Ч Центромера ¦ ¦ Щ Д I
4-
\
\
\
\
центромеры VI аутосомной хромосомы (схема 6) и занимает расстояние около 2 сантиморган между генами, кодирующими гипоксолазу (GL0) и мочевой пепсиноген 5 (Pg5). В пределах системы HLA могут разместиться около 105—106 генов.
Одна из характерных особенностей системы HLA — это высокий полиморфизм. Первый антиген был открыт Ж. Доссе в 1954 г., а к настоящему времени их более 100. Комитетом ВОЗ (1984) признаны следующие локусы системы HLA: HLA — А (23 антигена), HLA — В (49), HLA — С (8), HLA — DR (16), HLA — D (19), HLA — DQ (3), HLA — DP (6).
Гены, кодирующие антигены системы HLA, принято делить на 4 класса. К 1-му относят локусы А, В и С, ко 2-му — структуры D-области, в которых в свою очередь выделяют 4 сублокуса: HLA — D, HLA — DR, HLA — DQ и HLA — DP. В состав 3-го класса входят гены компонентов комплемента С2, С4а, С4ь, пропердиновый фактор Bf. Наконец, к 4-му классу условно отнесены гены, связь которых с системой HLA еще нуждается в доказательствах.
В биохимическом отношении HLA-антигены 1-го и 2-го классов состоят из двух почти равных гликополипептидных цепей а с относительной молекулярной массой 33—35 и цепей р с относительной молекулярной массой 28—31. Каждая цепь состоит из двух наружных доменов (ai, а: и Pi, р2 соответственно), имеет трансмембранную гидрофобную часть и цитоплазматический участок цепи. Различия аминокислотных последовательностей, характерные для определенных антигенов HLA, сосредоточены в доменах oil и Pi, в то время как домены 0.2 и р2 обладают более постоянным аминокислотным составом
и имеют выраженную гомологию с Рг-микроглобулином. Эти черты строения сходны со структурными особенностями иммуноглобулинов, что дает основание ряду исследователей постулировать общность эволюционного генеза иммуноглобулинов и антигенов системы HLA. При этом распознавательная функция последних считается эволюционно более древней, нежели распознавание с помощью антител.
Антигены гистосовместимости 1-го класса HLA—А, В и С присутствуют практически во всех клетках организма, за исключением ранних эмбриональных клеток — нормальных и злокачественных. При этом их количество и степень экспрессии сильно варьируют в зависимости от типа ткани. В наибольшем количестве они представлены на лимфоцитах, клетках эпителия и эндотелия. Антигены локусов 1-го класса (HLA—А, В, С) занимают около 1% клеточной поверхности. При этом число молекул одной специфичности приближается к 7000 и достаточно постоянно, поскольку через 6—7 ч после их удаления наблюдается практически их полное численное восстановление.
Основной метод определения антигенов 1-го класса — «компле- ментзависимая цитотоксичность». На первом этапе к типируемым лимфоцитам добавляются иммунная сыворотка определенной специфичности, а затем комплемент. Если сыворотка «узнает» наличие конкретного антигена HLA на лимфоцитах, то затем с помощью комплемента вызывает их лизис, который может быть обнаружен, например, с помощью прижизненных красителей (трипанового голубого).
Говоря о биологической функции антигенов 1-го класса, следует отметить, что именно они главным образом выступают в качестве рецепторов для чужеродных антигенов. При этом, учитывая теорию двойного распознавания [Zinkernagel R., Doherty P., 1974], суть которой состоит в том, что агент распознается Т-клеткой как чужеродный лишь после того, как он соединится с собственным антигеном презен- тирующей клетки, значение антигенов HLA 1-го класса представляется чрезвычайно важным.
По данным J. Hildreth, A. McMichael (1981), вирус гриппа А может непосредственно взаимодействовать с HLA—D-антигенами. С другой стороны, Ж. Доссе (1981) полагает, что МНС обеспечивает взаимодействие не только между иммунокомпетентными, но и любыми другими клетками организма, обусловливая функциональное единство организма. При этом главную роль от отводит антигенам 1-го класса, тогда как антигены 2-го класса, с его точки зрения, выполняют функцию распознавания лишь в системе иммунокомпетентных клеток.
В процессе иммунного ответа антигенам 1-го класса принадлежит ведущая роль (функция рестрикции иммунного ответа) во взаимодействии между клеткой-эффектором (Т-киллером) и клеткой-«мишенью». С другой стороны, Т-хелперы, как предшественники, так и зрелые клетки, распознают антигены 2-го класса, результатом чего является синтез интерлейкина-2 (IL-2), без которого не происходит созревания предшественника Т-киллеров — образование зрелых эффекторных кле
ток. На самом деле ситуация, по-видимому, еще сложнее. Е. Ball и P. Stastny (1982) при иммунохимическом анализе срезов отторгнутых аллотрансплантатов почки представили доказательства того, что Т- хелперы могут in vivo выполнять функцию, которая ранее приписывалась только киллерам. При этом взаимодействуют они с антигенами 2-го класса, которые широко представлены на эндотелии гломерул почек.
Таким образом, прослеживается тесная взаимосвязь антигенов 1-го и 2-го классов в процессе иммунного ответа.
Антигены 2-го класса региона D в последние годы являются предметом особо пристального внимания иммуногенетиков и иммунологов, поскольку именно с ними, как полагают, связаны гены иммунного ответа человека Ir (Immune response). Кроме того, они, по-видимому, играют ведущую роль во взаимодействии иммунокомпетентных клеток при развитии иммунного ответа. Существование локуса, отличного от локусов А, В и С, которые обнаруживаются в серологических цито- токсических реакциях, было установлено в реакции смешанных культур лимфоцитов MLC (Mixed Lymphocyte Culture). При этом лимфоциты от двух доноров, идентичных по МНС, в смешанной культуре не образуют бластов, а неидентичные — образуют.
Были установлены следующие факты: сибсы, неидентичные по SD (serological defined)-aHTHreHaM могли давать отрицательную реакцию MLC, т. е. не формировать бластов, и, напротив, лимфоциты от сибсов, идентичных по антигенам 1-го класса, давали положительную реакцию в MLC. Аналогичные результаты были получены при исследовании лимфоцитов не родственных лиц.
Таким образом, было постулировано наличие нового региона в системе HLA — D (и его локусов DR, D, DQ и DP). HLA — DR-антигены экспрессированы в основном на В-лимфоцитах и макрофагах, а также выявляются на эпидермальных, эндотелиальных клетках и сперматозоидах. На Т-клетках HLA — DR-антигены в обычном состоянии не обнаруживаются, но, будучи активированными in vitro и in vivo митогенами или аллогенными лимфоцитами, они экспрессируют антигены примерно на том же уровне, что и В-лимфоциты. Возможно, этот факт имеет определенный биологический смысл, ибо экспрессия генов ведет к взаимодействию клеток с развитием целой цепи последовательных событий и, возможно, как стимулирующий фактор, индуцирующий иммунную реакцию, она должна проявляться только при необходимости.
Один из важнейших разделов иммуногенетики — связь системы HLA и заболеваний. Этот аспект обстоятельно изложен в специальных обзорах и монографиях [Зарецкая Ю. М., 1983; Алексеев JI. П., 1985; Медуницин Н. В., Алексеев JI. П., 1987; Шебалин В. Н., Серова JI. Д., 1988]. Здесь лишь кратко будет упомянуто об основных этапах и направлениях поиска по данной проблеме. Первый (чисто феноменологический) этап начался, по-видимому, с изучения HLA-фенотипов лейкоцитов при болезни Ходжкина [Amiel J. L., 1967], после чего
последовало большое число публикаций, основной целью которых было установление достоверных взаимосвязей между антигенами системы HLA и различными заболеваниями, чтобы попытаться прогнозировать (сформировать группы риска) развитие заболевания на основании иммуногенетических исследований. При ряде заболеваний (синдром Рейтера, сальмонеллезный и иерсениозный артриты, дерматозы герпесной этиологии, гемохроматоз и некоторых др.) уже были выявлены сильные ассоциации с высоким риском развития болезни. Уникальным в этом отношении оказался анкилозирующий спондилит, связанный с HLA — В27. Его показатель относительного риска — около 100.
С конца 70-х годов стало развиваться новое направление — изучение механизмов связи между системой HLA и заболеваниями. L. Lamm (1979) выделил два вида ассоциаций: генетическую детерминированность и генетическую ассоциацию. Под первой из них понимается истинное сцепление «патологического гена» с антигенами системы HLA, при котором каждый раз «патологический ген» передается по наследству совместно с тем или иным локусом HLA. Такое явление представляет собой чрезвычайно ценный прогностический признак, однако встречается оно крайне редко (и для HLA не описано), например, дефицит фермента 2,1-гидроксилазы, гемохроматоз, абсолютная недостаточность С2- и С4-компонентов комплемента. Как правило, связь HLA и заболеваний проявляется в форме ассоциаций. Относительно их механизмов предложено несколько гипотез, каждая из которых имеет экспериментальные и клинические обоснования.
Первая из них — рецепторная гипотеза — предлагает рассматривать HLA-антигены как рецепторы, к которым прикрепляются вирусы, что облегчает проникновение последних в клетку и инициацию патологического процесса. Позднее справедливость этого предположения была доказана не только для вируса, но и для ряда химических и биологических препаратов. В частности, J. U. J. Glass и соавт. (1982) показали, что обработка пенициллином клеток, имеющих на своей поверхности антигены HLA — А9, Bw35 и В15, блокирует распознавание этих клеток соответствующими анти-Ш,А-сыворотками.
Гипотеза молекулярной мимикрии основана на структурном сходстве HLA-антигенов и антигенов некоторых болезнетворных агентов (микробов или вирусов). Вследствие этого организм, воспринимая такие антигены как свои, остается толерантным к ним. Гипотеза модификаций HLA-антигенов исходит из предпосылки о том, что вирус или другой инфекционный или неинфекционный агент может изменять структуру своих антигенов, после чего они распознаются в организме как чужеродные и индуцируют аутоиммунный процесс.
Н. В. Медуницин, J1. П. Алексеев (1987) полагают, что одним из механизмов ассоциации предрасположенности к определенным заболеваниям с генами D-региона является нарушение функции последнего в обеспечении «нормального» взаимодействия между иммунокомпе-
тентными клетками. С их точки зрения, нарушение именно этой функции имеет место при* первичных иммунодефицитах.
Одним из интереснейших направлений явилось изучение силы иммунного ответа на различные антигены в зависимости от HLA-фенотипа. В частности, было показано [Mann D. et al., 1985], что у больных, леченных инсулином, высокий уровень пролиферативного ответа на бычий инсулин ассоциировался с антигеном HLA — DP4, а на свиной — с DP3. Установлена также связь между экспрессией HLA — Al, В8, DP3 в составе фенотипа и повышенной иммунологической реактивностью. Показано, с другой стороны, что как у здоровых лиц, в фенотипе которых обнаруживался HLA — DP3, так и больных с аутоиммунными и аллергическими процессами — носителей антигенов HLA — В8, DP3 снижена активность Т-супрессоров, вследствие чего имеет место высокий иммунный ответ на стимуляцию многими антигенами.
С этими же антигенами ассоциирована высокая активность клеток в смешанной культуре аутологичных лимфоцитов, которая является признаком аутоиммунного процесса. Выявлено, что наличие в фенотипе антигенов HLA — В7, DP2 сочетается с повышенной супрессорной активностью [Gyodi Е. et al., 1982] и пониженной активностью естественных киллеров. Например, у больных системной красной волчанкой (в ее патогенезе ведущую роль играет снижение супрессорной функции), в фенотипе которых представлены антигены HLA — В7, DR2, угнетение супрессорных функций выражено слабее, а прогноз заболевания лучше [Rigby Р. О., 1978].
Несмотря на интенсивное развитие работ по изучению функций МНС, многое в этом вопросе остается неясным. В частности, мало изучены связи HLA-системы с функциональным состоянием ряда других систем иммунитета.
Мы попытались установить взаимодействие между антигенами системы HLA (локусов А и В) и функциональным состоянием отдельных звеньев иммунитета у здоровых и больных различными формами пневмонии детей. Идентификация HLA-антигенов проводилась методом комплементзависимой цитотоксичности по Ю. М. Зарецкой (1983). Иммунный статус исследовался по следующим показателям: спонтанные ЕРОК (Т-клетки), теофиллинчувствительные (обогащенные Т-хел- перами) и теофиллинрезистентные (обогащенные Т-супрессорами) РОК, ЕАС—РОК (В-клетки), «0»-клетки, РБТЛ с ФГА, уровни IgG, IgA и IgM, фагоцитарное число и фагоцитарный индекс.
Были обследованы 181 здоровый ребенок в возрасте от 1 мес до 3 лет и 160 детей, больных различными формами пневмонии (все обследованные азербайджанской национальности). Сравнивалась частота встречаемости отдельных антигенов в группе здоровых и больных детей с тем, чтобы выяснить, нет ли доминирования каких-либо из них у больных пневмонией и нельзя ли на этой основе формировать группы риска. Оказалось, что по частоте встречаемости в обеих сравниваемых группах первые 3 мес занимают одни и те же антигены (А2, В35, АЗ). Далее их порядок меняется, однако несущественно. Макси
мальные показатели для В8 у 16% здоровых детей и 21% — у больных, т. е. антигена, существенно превалирующего у больных детей, не обнаружено.
На втором этапе была предпринята попытка установить, существует ли связь между какими-либо антигенами системы HLA и уровнем показателей тех или иных звеньев иммунной системы. С этой целью отбирались дети с конкретным HLA-антигеном в гаплотипе и вычислялись средние значения всех исследованных иммунологических параметров (обозначены как «среднее +»). Затем отбирались дети без данного антигена и также вычислялись средние значения всех иммунологических показателей («среднее -»). Сопоставлялись «среднее +» и «среднее -» для каждого иммунологического показателя и при наличии статистически достоверных различий считалось, что данный антиген связан с количественным проявлением (высоким и низким) того или иного иммунологического параметра. Иными словами, если «среднее +» (группа детей, у которых присутствует исследуемый антиген), было достоверно выше, чем «среднее -», то антиген связан (или коррелирует) с сильным проявлением иммунологического параметра, если ниже, то со слабой функцией исследуемого звена иммунитела.
Таким образом, было выявлено 7 из 37 обследованных антигенов HLA, присутствие которых достоверно связано с сильным или слабым проявлением функции того или иного иммунологического параметра. Некоторые из них оказались связанными с противоположными функциональными состояниями одной и той же системы. Так, HLA — А1 связан с сильным проявлением функции Т-системы иммунитета (высокие значения в реакции Е-РОК), тогда как HLA — В15 — со слабым.
Проведенная работа представляет собой традиционный подход к поиску взаимосвязей между антигенами системы HLA и различными заболеваниями. Была предпринята попытка углубить анализ и поставлена задача определить, какими будут заначения иммунологического параметра, если антигены HLA, связанные с противоположными проявлениями функции, встретятся у одного ребенка. Проявится ли доминирование какого-либо антигена или влияние их будет взаимно нивелировано. Результаты оказались неожиданными: антигены А1 и В15, связанные соответственно с высокими и низкими значениями Е-РОК, а также В8 и В15, связанные с низкими и высокими показателями ЕАС-РОК, ни разу (из 34 возможных теоретических случаев) не встретились в пределах одного фенотипа у здоровых детей. Антигены же, сочетающиеся с однонаправленными функциями одного и того же иммунологического звена, встречались в пределах одного фенотипа: Al, В8 — в 12 случаях и 29 теоретически возможных; В8, В12 — в 8 случаях из 23.
Поскольку у здоровых детей выявилась столь неожиданная закономерность, было решено проверить, сохраняется ли она у больных. Оказалось, что сочетание антигенов, «кодирующих» противоположно направленные функции, в группе больных детей встретилось в двух из 64 теоретически возможных случаев, тогда как сочетание антигенов,
связанных с однонаправленными изменениями функции, встретилось в 34 случаях из 65 возможных (52%). Таким образом, как в группе здоровых, так и среди больных детей выявлена своеобразная закономерность. И хотя на основании имеющегося числа наблюдений (341 ребенок) нельзя сделать категорические выводы, однако полученные данные свидетельствуют о том, что, вероятно, есть какие-то ограничения (рестрикция) для объединения в одном фенотипе антигенов, связанных с противоположными функциональными состояниями одного и того же иммунологического звена.
Важным является вопрос, ассоциированы ли одни и те же антигены HLA — А и В с одними и теми же иммунологическими показателями у здоровых и больных детей. Если это так, то определяют ли они одну и ту же функциональную направленность отдельных звеньев иммуни- тела, т. е. низкие или высокие значения исследуемых параметров?
При анализе полученных данных выяснилось, что ни в одном случае в сравниваемых группах здоровых и больных детей нет совпадения, достоверной связи того или иного антигена системы HLA с тем или иным иммунологическим параметром. В частности, у здоровых детей антиген HLA — А1 достоверно связан с высокими значениями Е-РОК и РБТЛ и встречается часто (у 35 из 181—19,3%), тогда как у больных этот антиген при практически такой же частоте встречаемости (18,75%) достоверно не связан ни с одним иммунологическим параметром.
Таким образом, в ходе иммунологических исследований не удалось получить «опорных» для клиники показателей, которые можно было бы практически использовать для диагностики, прогноза и формирования групп риска. Однако были обнаружены интересные и в значительной мере неожиданные закономерности, которым сегодня нельзя дать удовлетворительного объяснения и лишь можно ограничиться феноменологической констатацией. Во всяком случае выявленные особенности, безусловно, подлежат дальнейшему углубленному изучению.
Таким образом, МНС представляет собой большую полиморфную систему генов и их представителей (антигенов) на мембранах практически всех клеток организма, участвующих в ряде важнейших биологических процессов: в распознавании антигенами МНС бактерий и вирусов, в реакции на трансплантационные антигены, во взаимодействии иммунокомпетентных клеток в ходе развития клеточного и гуморального иммунного ответа, в генетическом контроле активности системы комплемента. Кроме того, МНС осуществляет генетический контроль за феноменами, не относящимися прямо к функции иммунитета (уровень некоторых гормонов, процесс размножения и развития, взаимодействие между любыми, а не только иммунокомпетент- ными клетками).
По-видимому, следует согласиться с теми авторами, которые считают функцию генетического контроля со стороны МНС за общебиологическими процессами важнейшей и первичной, тогда как генетический контроль иммунного ответа — вторичной и производной.
А так же в разделе «ГЛАВА 3 СИСТЕМА HLA И ИММУНОЛОГИЧЕСКАЯ РЕАКТИВНОСТЬ »
- СПИСОК СОКРАЩЕНИЙ
- ПРЕДИСЛОВИЕ
- ВВЕДЕНИЕ
- ГЛАВА 1 ГУМОРАЛЬНЫЕ ФАКТОРЫ ИММУНИТЕТА
- Острофазный ответ организма
- Система комплемента
- Интерферон (интерфероны)
- ГЛАВА 2 КЛЕТОЧНЫЕ ОСНОВЫ ИММУННЫХ РЕАКЦИЙ
- Медиаторы клеточного иммунитета
- Нейтрофилы
- СТАНОВЛЕНИЕ И РАЗВИТИЕ ИММУННОЙ СИСТЕМЫ В ОНТОГЕНЕЗЕ
- ГЛАВА 5 ИММУНОДЕФИЦИТНЫЕ состояния
- Наследственные (первичные) формы ИДС
- Лечение ИДС
- Рецидивирующие и хронические заболевания легких