
На первом этапе непосредственного исследования структуры генома человека, когда еще не существовала методология генной инженерии, для изучения ДНК применяли традиционные физико–химические методы. В этих опытах использовали суммарные препараты ДНК, целиком выделенные из ядер клеток человека.
Пожалуй, первые сведения о молекулярной структуре генома человека были получены в результате центрифугирования в пробирке растворов ДНК в хлористом цезии при довольно высоких скоростях. В процессе вращения соли цезия создают тонкие слои раствора с различной плотностью (градиент плотности) вдоль пробирки и молекулы ДНК перемещаются в этом градиенте, пока не достигнут такой области, где плотность солевого раствора будет точно такой же, как их собственная. А плотность ДНК сильно зависит от содержания АТ–и ГЦ–пар нуклеотидов, т. е., как говорят, от нуклеотидного состава. Оказалось, что основная масса ДНК человека после центрифугирования располагается преимущественно в одной зоне градиента (это соответствует среднему содержанию ГЦ–пар в геноме человека, равному 42%). Однако наряду с этим неожиданно обнаруживались и небольшие (минорные) полосы, в которых также содержались молекулы ДНК, но с иной плотностью и, следовательно, с иным содержанием нуклеотидных пар. Такие минорные, или дополнительные фракции ДНК получили название «сателлитных». Такое имя дали этим фракциям не случайно. В то время как раз был запущен первый советский спутник (лат. satellitis – спутник). Это и натолкнуло исследователей на такое название.
Вскоре после того, как была установлена двухспиральная структура ДНК, обнаружили, что при сильном нагревании ДНК две ее цепи расходятся (расплавляются) и ДНК из двунитевой превращается в однонитевую. Это приводит к нарушению ее естественной структуры, что получило отражение в названии данного процесса – денатурация ДНК. Однако при охлаждении денатурированной ДНК комплементарные цепи находят друг друга и соединяются строго так, как они располагались в исходной неденатурированной молекуле, по типу застежки – «молнии». Этот процесс получил название рена–турации или реассоциации. Сразу же после обнаружения этого явления оно было использовано экспериментаторами в целях изучения структуры генома.
Немного совсем простой математики для тех самых любопытных, кто недавно закончил ВУЗ, изучал химию и еще не забыл все, чему его учили. Процесс реассоциации ДНК во многом сходен с обычной химической реакцией второго порядка и, по этой причине, может быть описан довольно простой формулой:
Сt/C0, = 1/(1 + k2(C0t)),
где C0 и Сt – концентрации однонитевых ДНК соответственно в начальный (нулевой) момент времени и в момент времени t после начала реакции реассоциации, k2 – константа скорости реакции второго порядка.
Для удобства изображения процесса реассоциации строят график, в котором по оси ординат откладывают долю реассоциированных молекул, а по оси абсцисс – величину C0t. При таком изображении кривые кинетики реассоциации ДНК простейших организмов (вирусов и бактерий) имеют S – образный вид. Это соответствует кинетике химической реакции второго порядка. Если в эксперименте наблюдают отклонения от этой кривой, то это должно указывать на гетерогенность цепей ДНК по скорости взаимодействия друг с другом: одни реагирует быстрее, другие, соответственно, медленнее. Когда ДНК человека порезали на куски небольшого размера и измерили кинетику их реассоциации, то обнаружилось, что кривая, описывающая этот процесс, далека от стандартной (рис. 12).
Это явилось результатом того, что в ДНК человека имеются нук–леотидные последовательности, реассоциирующие с разной скоростью, а суммарная кривая реакции, наблюдаемая в опыте, отражает совокупность множества независимых реакций второго порядка. Когда были проведены эти эксперименты, уже существовал математический аппарат, позволяющий, хотя и очень грубо, вычленять из сложной суммарной кривой отдельные относительно однородные кинетические компоненты. Различия в скоростях реассоциации разных компонентов ДНК человека были связаны с разной представленностью в ДНК отдельных нуклеотидных последовательностей. Участки, которые присутствуют в геноме всего один раз, назвали уникальными. Если в геноме определенная нуклеотидная последовательность не уникальна, а представлена неким числом одинаковых копий (т. е. повторяется, отсюда и название – повторяющаяся), то такая последовательность, естественно, по чисто химическим законам, будет в растворе находить комплементарную цепь и взаимодействовать с ней (реассоциировать) значительно быстрее первой.
В результате такого анализа нашли, что в геноме человека около 75% участков ДНК представлены 1 копией (уникальные) на гаплоидный геном (естественно, в ядре, являющемся диплоидным, имеется 2 копии каждой уникальной нуклеотидной последовательности). Остальную часть генома составляют повторяющиеся последовательности (повторы), среди которых 10% представлены очень быстро реассоциирующими повторами (104 и более копий на геном) и около 15% – умеренными повторами. Заметим сразу, что в дальнейшем эти оценки были существенно пересмотрены. Однако некоторые из этих результатов остались в силе до сих пор.
Первоначальные анализы показали, что среди быстро взаимодействующих друг с другом при реассоциации фрагментов ДНК присутствует некоторое количество таких, кинетика реассоциации которых отличается от реакции второго порядка. Причина этому оказалось в том, что эти участки представляют собой так называемые обращенные повторы, или палиндромы – взаимокомплементарные последовательности, расположенные не на разных, а на одной нити ДНК.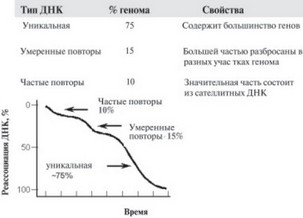
Рис. 12. Кинетика восстановления двунитевых молекул из искусственно разделенных комплементарных цепей ДНК человека (реассоциация). ДНК разбивают на небольшие фрагменты, денатурируют путем нагревания, а затем при охлаждении разошедшиеся цепи ДНК вновь соединяются. Чем чаще в смеси встречаются те или иные последовательности, тем быстрее они находят друг друга в растворе и реассоциируют. По этой кинетике определяли общее содержание повторяющихся и неповторяющихся (уникальных) нуклеотидных последовательностей в геноме человека
Таким образом, в ДНКовом текте присутствуют «предложения» – палиндромы («перевертыши»), одинаково читаемые слева направо и справа налево. Перевертыши хорошо известны из литературы – это предложения, которые читаются одинаково слева направо и справа налево без учета знаков препинания и интервалов между словами. В качестве примера приведем один из таких перевертышей:
УЖРЕДКОРУКОЮОКУРОКДЕРЖУ.
В ДНК перевертышами называют отрезки двойной антипараллельной спирали, которые имеют одинаковую нуклеотидную последовательность при чтении по обеим цепям в одинаковом направлении. Это выглядит как, например, в ниже приведенном случае:

Здесь стрелками показано направление цепей ДНК, а звездочками – водородные связи, образуемые между парами нуклеотидов. Общее число таких «перевертышей» в геноме человека оценено в интервале от 105 до 106 . При этом они относительно равномерно распределены по ДНК.
Имеются в геноме человека и нуклеотидные последовательности, которые на всем своем протяжении построены из одной единственной «буквы». Если в одной цепи ДНК эта буква А, то в другой цепи, соответственно, будет буква Т. Такие участки названы гомополимер–ными. В случае приведенного выше примера последовательность нуклеотидов записывается, как поли(А) – поли(Т). Таким образом, выяснилось, что в геноме человека (а параллельно это изучали и в геномах других организмов) имеются все варианты нуклеотидных последовательностей, состоящих из 4 «букв», которые только можно себе мысленно представить.
Используя различные модификации метода реассоциации ДНК, установили также, что в большей части генома человека повторяющиеся и уникальные нуклеотидные последовательности перемежаются друг с другом, а средние длины перемежающихся повторяющихся и уникальных фрагментов ДНК составляют соответственно 300 и 2000 п. н.
Приведенные выше данные были, конечно же, усредненными и поверхностно отражали общую картину устройства генома человека. Тем не менее они послужили хорошей основой для дальнейших более углубленных исследований. Важно, что одновременно такие же работы проводились и с использованием других эукариотичес–ких организмов. Многие моменты оказались сходными у разных высших организмов и растений. Так постепенно начали вырисовываться в общих чертах основные принципы организации генома человека.
Пожалуй, первые сведения о молекулярной структуре генома человека были получены в результате центрифугирования в пробирке растворов ДНК в хлористом цезии при довольно высоких скоростях. В процессе вращения соли цезия создают тонкие слои раствора с различной плотностью (градиент плотности) вдоль пробирки и молекулы ДНК перемещаются в этом градиенте, пока не достигнут такой области, где плотность солевого раствора будет точно такой же, как их собственная. А плотность ДНК сильно зависит от содержания АТ–и ГЦ–пар нуклеотидов, т. е., как говорят, от нуклеотидного состава. Оказалось, что основная масса ДНК человека после центрифугирования располагается преимущественно в одной зоне градиента (это соответствует среднему содержанию ГЦ–пар в геноме человека, равному 42%). Однако наряду с этим неожиданно обнаруживались и небольшие (минорные) полосы, в которых также содержались молекулы ДНК, но с иной плотностью и, следовательно, с иным содержанием нуклеотидных пар. Такие минорные, или дополнительные фракции ДНК получили название «сателлитных». Такое имя дали этим фракциям не случайно. В то время как раз был запущен первый советский спутник (лат. satellitis – спутник). Это и натолкнуло исследователей на такое название.
Вскоре после того, как была установлена двухспиральная структура ДНК, обнаружили, что при сильном нагревании ДНК две ее цепи расходятся (расплавляются) и ДНК из двунитевой превращается в однонитевую. Это приводит к нарушению ее естественной структуры, что получило отражение в названии данного процесса – денатурация ДНК. Однако при охлаждении денатурированной ДНК комплементарные цепи находят друг друга и соединяются строго так, как они располагались в исходной неденатурированной молекуле, по типу застежки – «молнии». Этот процесс получил название рена–турации или реассоциации. Сразу же после обнаружения этого явления оно было использовано экспериментаторами в целях изучения структуры генома.
Немного совсем простой математики для тех самых любопытных, кто недавно закончил ВУЗ, изучал химию и еще не забыл все, чему его учили. Процесс реассоциации ДНК во многом сходен с обычной химической реакцией второго порядка и, по этой причине, может быть описан довольно простой формулой:
Сt/C0, = 1/(1 + k2(C0t)),
где C0 и Сt – концентрации однонитевых ДНК соответственно в начальный (нулевой) момент времени и в момент времени t после начала реакции реассоциации, k2 – константа скорости реакции второго порядка.
Для удобства изображения процесса реассоциации строят график, в котором по оси ординат откладывают долю реассоциированных молекул, а по оси абсцисс – величину C0t. При таком изображении кривые кинетики реассоциации ДНК простейших организмов (вирусов и бактерий) имеют S – образный вид. Это соответствует кинетике химической реакции второго порядка. Если в эксперименте наблюдают отклонения от этой кривой, то это должно указывать на гетерогенность цепей ДНК по скорости взаимодействия друг с другом: одни реагирует быстрее, другие, соответственно, медленнее. Когда ДНК человека порезали на куски небольшого размера и измерили кинетику их реассоциации, то обнаружилось, что кривая, описывающая этот процесс, далека от стандартной (рис. 12).
Это явилось результатом того, что в ДНК человека имеются нук–леотидные последовательности, реассоциирующие с разной скоростью, а суммарная кривая реакции, наблюдаемая в опыте, отражает совокупность множества независимых реакций второго порядка. Когда были проведены эти эксперименты, уже существовал математический аппарат, позволяющий, хотя и очень грубо, вычленять из сложной суммарной кривой отдельные относительно однородные кинетические компоненты. Различия в скоростях реассоциации разных компонентов ДНК человека были связаны с разной представленностью в ДНК отдельных нуклеотидных последовательностей. Участки, которые присутствуют в геноме всего один раз, назвали уникальными. Если в геноме определенная нуклеотидная последовательность не уникальна, а представлена неким числом одинаковых копий (т. е. повторяется, отсюда и название – повторяющаяся), то такая последовательность, естественно, по чисто химическим законам, будет в растворе находить комплементарную цепь и взаимодействовать с ней (реассоциировать) значительно быстрее первой.
В результате такого анализа нашли, что в геноме человека около 75% участков ДНК представлены 1 копией (уникальные) на гаплоидный геном (естественно, в ядре, являющемся диплоидным, имеется 2 копии каждой уникальной нуклеотидной последовательности). Остальную часть генома составляют повторяющиеся последовательности (повторы), среди которых 10% представлены очень быстро реассоциирующими повторами (104 и более копий на геном) и около 15% – умеренными повторами. Заметим сразу, что в дальнейшем эти оценки были существенно пересмотрены. Однако некоторые из этих результатов остались в силе до сих пор.
Первоначальные анализы показали, что среди быстро взаимодействующих друг с другом при реассоциации фрагментов ДНК присутствует некоторое количество таких, кинетика реассоциации которых отличается от реакции второго порядка. Причина этому оказалось в том, что эти участки представляют собой так называемые обращенные повторы, или палиндромы – взаимокомплементарные последовательности, расположенные не на разных, а на одной нити ДНК.
Рис. 12. Кинетика восстановления двунитевых молекул из искусственно разделенных комплементарных цепей ДНК человека (реассоциация). ДНК разбивают на небольшие фрагменты, денатурируют путем нагревания, а затем при охлаждении разошедшиеся цепи ДНК вновь соединяются. Чем чаще в смеси встречаются те или иные последовательности, тем быстрее они находят друг друга в растворе и реассоциируют. По этой кинетике определяли общее содержание повторяющихся и неповторяющихся (уникальных) нуклеотидных последовательностей в геноме человека
Таким образом, в ДНКовом текте присутствуют «предложения» – палиндромы («перевертыши»), одинаково читаемые слева направо и справа налево. Перевертыши хорошо известны из литературы – это предложения, которые читаются одинаково слева направо и справа налево без учета знаков препинания и интервалов между словами. В качестве примера приведем один из таких перевертышей:
УЖРЕДКОРУКОЮОКУРОКДЕРЖУ.
В ДНК перевертышами называют отрезки двойной антипараллельной спирали, которые имеют одинаковую нуклеотидную последовательность при чтении по обеим цепям в одинаковом направлении. Это выглядит как, например, в ниже приведенном случае:
Здесь стрелками показано направление цепей ДНК, а звездочками – водородные связи, образуемые между парами нуклеотидов. Общее число таких «перевертышей» в геноме человека оценено в интервале от 105 до 106 . При этом они относительно равномерно распределены по ДНК.
Имеются в геноме человека и нуклеотидные последовательности, которые на всем своем протяжении построены из одной единственной «буквы». Если в одной цепи ДНК эта буква А, то в другой цепи, соответственно, будет буква Т. Такие участки названы гомополимер–ными. В случае приведенного выше примера последовательность нуклеотидов записывается, как поли(А) – поли(Т). Таким образом, выяснилось, что в геноме человека (а параллельно это изучали и в геномах других организмов) имеются все варианты нуклеотидных последовательностей, состоящих из 4 «букв», которые только можно себе мысленно представить.
Используя различные модификации метода реассоциации ДНК, установили также, что в большей части генома человека повторяющиеся и уникальные нуклеотидные последовательности перемежаются друг с другом, а средние длины перемежающихся повторяющихся и уникальных фрагментов ДНК составляют соответственно 300 и 2000 п. н.
Приведенные выше данные были, конечно же, усредненными и поверхностно отражали общую картину устройства генома человека. Тем не менее они послужили хорошей основой для дальнейших более углубленных исследований. Важно, что одновременно такие же работы проводились и с использованием других эукариотичес–ких организмов. Многие моменты оказались сходными у разных высших организмов и растений. Так постепенно начали вырисовываться в общих чертах основные принципы организации генома человека.