
Успехи в изучении генома человека, а еще ранее – проведенный анализ отдельных генов других высших организмов, привели к ряду принципиально новых и совершенно неожиданных для генетиков результатов в отношении устройства генов, особенно тех, которые кодируют белки. Со времени появления центральной догмы молекулярной биологии думали, что непрерывная линейная последовательность нуклеотидов в ДНК с помощью триплетов нуклеоти–дов кодирует непрерывную последовательность нуклеотидов в мРНК, а последняя как посредник передает эту информацию на рибосомы, которые, считывая ее, осуществляют синтез линейных молекул белков. Это соответствовало ситуации, которую первоначально наблюдали при изучении генов у бактерий. Однако у высших организмов, включая человека, (и даже у некоторых низших) картина оказалась более сложной.
Когда сравнивали мРНК и соответствующий белок, то действительно обнаруживали непрерывность их нуклеотидных и аминокислотных последовательностей. То же наблюдали и при сопоставлении последовательностей аминокислот в белках и нуклеотидов в геномах вирусов. Однако с появлением возможности секвенировать протяженные участки ДНК (т. е. непосредственно генов) как у некоторых вирусов, так и у высших организмов получили совершенно неожиданные результаты. Внутри нуклеотидных последовательностей ДНК, кодирующих белок, были обнаружены какие–то иные, ничего не кодирующие участки. Чудеса, да и только! В чем же дело? Оказалось, что многие гены «расчленены» на отдельные куски. Одни из этих кусков, как и положено, кодируют белок (их назвали экзонами), а другие никакие белки кодировать не способны (их назвали интронами) и расположены между экзонами. Таким образом, в гене кодирующие и не кодирующие участки перемежаются друг с другом (рис. 18). Так устроено большинство белок–кодирующих генов. Хотя имеются и исключения. В этих случаях говорят, что ген устроен из одного экзона и интронов не содержит.
Такое разорванное (экзон–интронное) строение оказалось характерным для подавляющего большинства (хотя и не для всех) генов человека. Вместе с тем у большинства простейших организмов инт
Рис. 18. Схематическое изображение экзон–интронного строения некоторых генов человека. Экзоны – прямоугольнники, интроны – линии между прямоугольниками. Темные прямоугольники – участки генов, кодирующие белок, белые прямоугольники в первом и последнем экзонах – специальные участки гена, кодирующие мРНК, но не кодирующие белок
роны не обнаружены. Размеры интронов часто значительно превышают размеры экзонов, что существенно увеличивает общую длину нуклеотидных последовательностей ДНК, образующих ген. При этом в геноме человека интроны в целом существенно длиннее, чем у других организмов.
Каким же образом разорванный ДНКовый текст реализуется в клетках в виде неразорванного РНКового, а затем белкового? Выяснилось, что первоначально при транскрипции гена синтезируется большой РНК–предшественник, копия ДНКового текста (с экзона–ми и интронами вместе). Далее в результате работы специфических ферментов происходит нарезание РНК–предшественника на куски. Те из них, которые ничего не кодируют (интроны), отбрасываются, а кодирующие куски (экзоны) соединяются между собой, обычно в том же порядке, как они располагались исходно в ДНК. В результате формируется зрелая молекула мРНК. Этот сложный процесс и получил название сплайсинга (рис. 19).
В интронах содержатся особые сигналы, которые узнаются специальными ферментами, осуществляющими сплайсинг. Так, в 98% случаев интроны начинаются с динуклеотида ГТ, а заканчиваются динуклеотидом АГ. Они–то и служат главными сигналами для правильного осуществления сплайсинга.
Во всем этом и состоит основное ноу–хау в организации генов человека и других эукариотических генов. «Лоскутное» устройство большинства генов – чрезвычайно важное эволюционное приобретение высших организмов. За счет сплайсинга в РНК может происходить соединение не только между соседними экзонными нуклео–тидными последовательностями, но и между другими, отстоящими порой в гене на значительном расстоянии. Это называют альтернативным сплайсингом (рис. 20). В геноме человека альтернативный сплайсинг характерен для более трети генов. Этому можно привести следующую аналогию: из слова «администрация» за счет удаления отдельных букв и слогов можно образовать множество совершенно разных по смыслу слов: ад, министр, амнистия, астра, нация. Существование множества альтернативных вариантов сплайсинга в конечном итоге обеспечивает появление более чем одного белка при экспрессии одного единственного гена. В частности, на гене белка тро–понина человека, содержащем 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных продукта! Сейчас подсчитано, что в среднем один ген у человека способен кодировать около 3–х разных белков. Альтернативный сплайсинг зависит от множества внутриклеточных и внешних факторов, в том числе, как показано недавно, даже от вирусного воздействия на клетку.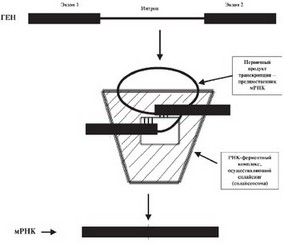
Рис. 19. Образование молекул мРНК на РНК–предшественнике в результате сплайсинга. Из РНК–предшественника вырезаются фрагменты, синтезированные с интронных последовательностей гена, а участки, синтезированные на экзонах, соединяются друг с другом, что в конечном итоге приводит к формированию зрелой функционально активной мРНК
Экзон–интронное строение генов дало принципиально новую возможность для эволюции генов – комбинировать отдельные элементы при формировании генов. В результате этого в эволюции возникали новые гены, кодирующие разные белки за счет разного сочетания экзонов одного и того же гена. То есть из уже готовых «слов» составляются совершенно разные «предложения». И такой подход, используемый природой, оказался весьма продуктивным.
Следует отметить, что и в зрелой мРНК, которая образуется уже после удаления из первичного транскрипта интронных нуклеотидных последовательностей, также не все участки кодируют белок. Однако, в отличие от интронов, такие последовательности нуклео–тидов расположены в начале и в конце молекулы (см. рис. 18). Их размеры также существенно различаются. Так, в разных мРНК чело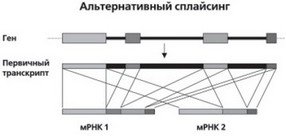
Рис. 20. Схематическое изображение альтернативного сплайсинга. Прямоугольниками разного оттенка обозначены экзоны, черными линиями – интроны. Показаны лишь две из множества возможных мРНК, образующихся на одном гене
века некодирующая область, расположенная в начале молекулы, имеет длину от 2800 до 18 нуклеотидов (в среднем 200 нуклеотидов), а та, которая расположена в конце, составляет от 8500 до 20 нуклеотидов (в среднем 1000 нуклеотидов). Эти некодирующие белок области гена очень важны для нормального функционирования мРНК, ее трансляции, стабильности и локализации в клетке. Как уже говорилось, на одном гене за счет альтернативного сплайсинга может образовываться несколько мРНК. Следует подчеркнуть, что и на одной уже сформировавшейся мРНК могут образовываться разные белки. Это происходит за счет наличия в 50% мРНК человека не одного, а двух АУГ–кодонов, являющихся старт–сигналами для синтеза белка на мРНК. За счет этого, в разных ситуациях клетка может начать синтез белка с разных мест мРНК, в результате чего образуются разные по длине, а порой и по своим свойствам, белки. Опять же пример из лингвистики. Если читать текст не с первой, а со второй буквы, смысл зачастую совершенно меняется (сравните: удочка и дочка).
В связи со всем этим понятно, что классическая формула генетиков «один ген – один белок», предложенная лауреатами Нобелевской премии Джоржем Бидлом и Эдуардом Татумом, оказывается в реальности справедливой лишь для небольшой группы генов, а многие гены (возможно, большинство) кодируют семейства родственных, но существенно отличающихся белков, то есть нередко действует принцип «один ген – много белков». За счет этого в геноме осуществляется принцип экономии генетического материала (компактная запись) и достигается большой набор генных продуктов – белков – на относительно небольшом числе генов.
В таблице 3 приведена общая характеристика белок–кодирующих генов, выявленных в результате секвенирования генома человека.
Таблица 3. Усредненные данные о структуре белок–кодирую–щих генов генома человека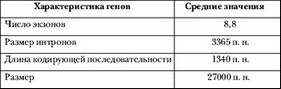
В реальности размеры разных генов, а также их экзонов и интро–нов сильно отличаются. И основной вклад в эту вариабельность вносят интроны. Уже говорилось, что максимальный по размерам ген дистрофина занимает в геноме человека 2,4 млн. п. н. Подавляющая часть занята именно интронами (их средний размер составляет несколько десятков тысяч п. н.). Наибольшую кодирующую последовательность в геноме человека имеет ген по имени титин. Его размер равен около 81000 п. н. Этот ген – чемпион и по числу интронов (178 штук!), и по длине единичного экзона (17106 п. н.).
Важно отметить, что разбивка генов на отдельные экзоны неслучайна. Индивидуальный экзон часто кодирует не просто какую–то часть белковой молекулы, а определенный фрагмент белка, выполняющий в целом белке отдельную функцию. Такой отдельный структурно–функциональный фрагмент, входящий в состав целого белка, называют доменом.
Когда сравнивали мРНК и соответствующий белок, то действительно обнаруживали непрерывность их нуклеотидных и аминокислотных последовательностей. То же наблюдали и при сопоставлении последовательностей аминокислот в белках и нуклеотидов в геномах вирусов. Однако с появлением возможности секвенировать протяженные участки ДНК (т. е. непосредственно генов) как у некоторых вирусов, так и у высших организмов получили совершенно неожиданные результаты. Внутри нуклеотидных последовательностей ДНК, кодирующих белок, были обнаружены какие–то иные, ничего не кодирующие участки. Чудеса, да и только! В чем же дело? Оказалось, что многие гены «расчленены» на отдельные куски. Одни из этих кусков, как и положено, кодируют белок (их назвали экзонами), а другие никакие белки кодировать не способны (их назвали интронами) и расположены между экзонами. Таким образом, в гене кодирующие и не кодирующие участки перемежаются друг с другом (рис. 18). Так устроено большинство белок–кодирующих генов. Хотя имеются и исключения. В этих случаях говорят, что ген устроен из одного экзона и интронов не содержит.
Такое разорванное (экзон–интронное) строение оказалось характерным для подавляющего большинства (хотя и не для всех) генов человека. Вместе с тем у большинства простейших организмов инт
Рис. 18. Схематическое изображение экзон–интронного строения некоторых генов человека. Экзоны – прямоугольнники, интроны – линии между прямоугольниками. Темные прямоугольники – участки генов, кодирующие белок, белые прямоугольники в первом и последнем экзонах – специальные участки гена, кодирующие мРНК, но не кодирующие белок
роны не обнаружены. Размеры интронов часто значительно превышают размеры экзонов, что существенно увеличивает общую длину нуклеотидных последовательностей ДНК, образующих ген. При этом в геноме человека интроны в целом существенно длиннее, чем у других организмов.
Каким же образом разорванный ДНКовый текст реализуется в клетках в виде неразорванного РНКового, а затем белкового? Выяснилось, что первоначально при транскрипции гена синтезируется большой РНК–предшественник, копия ДНКового текста (с экзона–ми и интронами вместе). Далее в результате работы специфических ферментов происходит нарезание РНК–предшественника на куски. Те из них, которые ничего не кодируют (интроны), отбрасываются, а кодирующие куски (экзоны) соединяются между собой, обычно в том же порядке, как они располагались исходно в ДНК. В результате формируется зрелая молекула мРНК. Этот сложный процесс и получил название сплайсинга (рис. 19).
В интронах содержатся особые сигналы, которые узнаются специальными ферментами, осуществляющими сплайсинг. Так, в 98% случаев интроны начинаются с динуклеотида ГТ, а заканчиваются динуклеотидом АГ. Они–то и служат главными сигналами для правильного осуществления сплайсинга.
Во всем этом и состоит основное ноу–хау в организации генов человека и других эукариотических генов. «Лоскутное» устройство большинства генов – чрезвычайно важное эволюционное приобретение высших организмов. За счет сплайсинга в РНК может происходить соединение не только между соседними экзонными нуклео–тидными последовательностями, но и между другими, отстоящими порой в гене на значительном расстоянии. Это называют альтернативным сплайсингом (рис. 20). В геноме человека альтернативный сплайсинг характерен для более трети генов. Этому можно привести следующую аналогию: из слова «администрация» за счет удаления отдельных букв и слогов можно образовать множество совершенно разных по смыслу слов: ад, министр, амнистия, астра, нация. Существование множества альтернативных вариантов сплайсинга в конечном итоге обеспечивает появление более чем одного белка при экспрессии одного единственного гена. В частности, на гене белка тро–понина человека, содержащем 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных продукта! Сейчас подсчитано, что в среднем один ген у человека способен кодировать около 3–х разных белков. Альтернативный сплайсинг зависит от множества внутриклеточных и внешних факторов, в том числе, как показано недавно, даже от вирусного воздействия на клетку.
Рис. 19. Образование молекул мРНК на РНК–предшественнике в результате сплайсинга. Из РНК–предшественника вырезаются фрагменты, синтезированные с интронных последовательностей гена, а участки, синтезированные на экзонах, соединяются друг с другом, что в конечном итоге приводит к формированию зрелой функционально активной мРНК
Экзон–интронное строение генов дало принципиально новую возможность для эволюции генов – комбинировать отдельные элементы при формировании генов. В результате этого в эволюции возникали новые гены, кодирующие разные белки за счет разного сочетания экзонов одного и того же гена. То есть из уже готовых «слов» составляются совершенно разные «предложения». И такой подход, используемый природой, оказался весьма продуктивным.
Следует отметить, что и в зрелой мРНК, которая образуется уже после удаления из первичного транскрипта интронных нуклеотидных последовательностей, также не все участки кодируют белок. Однако, в отличие от интронов, такие последовательности нуклео–тидов расположены в начале и в конце молекулы (см. рис. 18). Их размеры также существенно различаются. Так, в разных мРНК чело
Рис. 20. Схематическое изображение альтернативного сплайсинга. Прямоугольниками разного оттенка обозначены экзоны, черными линиями – интроны. Показаны лишь две из множества возможных мРНК, образующихся на одном гене
века некодирующая область, расположенная в начале молекулы, имеет длину от 2800 до 18 нуклеотидов (в среднем 200 нуклеотидов), а та, которая расположена в конце, составляет от 8500 до 20 нуклеотидов (в среднем 1000 нуклеотидов). Эти некодирующие белок области гена очень важны для нормального функционирования мРНК, ее трансляции, стабильности и локализации в клетке. Как уже говорилось, на одном гене за счет альтернативного сплайсинга может образовываться несколько мРНК. Следует подчеркнуть, что и на одной уже сформировавшейся мРНК могут образовываться разные белки. Это происходит за счет наличия в 50% мРНК человека не одного, а двух АУГ–кодонов, являющихся старт–сигналами для синтеза белка на мРНК. За счет этого, в разных ситуациях клетка может начать синтез белка с разных мест мРНК, в результате чего образуются разные по длине, а порой и по своим свойствам, белки. Опять же пример из лингвистики. Если читать текст не с первой, а со второй буквы, смысл зачастую совершенно меняется (сравните: удочка и дочка).
В связи со всем этим понятно, что классическая формула генетиков «один ген – один белок», предложенная лауреатами Нобелевской премии Джоржем Бидлом и Эдуардом Татумом, оказывается в реальности справедливой лишь для небольшой группы генов, а многие гены (возможно, большинство) кодируют семейства родственных, но существенно отличающихся белков, то есть нередко действует принцип «один ген – много белков». За счет этого в геноме осуществляется принцип экономии генетического материала (компактная запись) и достигается большой набор генных продуктов – белков – на относительно небольшом числе генов.
В таблице 3 приведена общая характеристика белок–кодирующих генов, выявленных в результате секвенирования генома человека.
Таблица 3. Усредненные данные о структуре белок–кодирую–щих генов генома человека
В реальности размеры разных генов, а также их экзонов и интро–нов сильно отличаются. И основной вклад в эту вариабельность вносят интроны. Уже говорилось, что максимальный по размерам ген дистрофина занимает в геноме человека 2,4 млн. п. н. Подавляющая часть занята именно интронами (их средний размер составляет несколько десятков тысяч п. н.). Наибольшую кодирующую последовательность в геноме человека имеет ген по имени титин. Его размер равен около 81000 п. н. Этот ген – чемпион и по числу интронов (178 штук!), и по длине единичного экзона (17106 п. н.).
Важно отметить, что разбивка генов на отдельные экзоны неслучайна. Индивидуальный экзон часто кодирует не просто какую–то часть белковой молекулы, а определенный фрагмент белка, выполняющий в целом белке отдельную функцию. Такой отдельный структурно–функциональный фрагмент, входящий в состав целого белка, называют доменом.