
Губки (Porifera). Наиболее просто организованные первичные многоклеточные — губки, клетки которых еще не объединены в ткани, уже имеют несколько морфологически и функционально самостоятельных клеточных типов. Среди них: пинакоциты — клетки, покрывающие внешнюю поверхность тела и стенки внутренних водоносных каналов, хоаноцнты — сферические или кубои- дальные, снабженные ресничками клетки, которые выстилают поверхность камер и обеспечивают движение воды в теле губок. Имеется также группа клеток, локализованных в мезоглее: колленциты и лофоциты, продуцирующие коллаген; спонгоциты, образующие спонгин; склероциты, ответственные за формирование игольчатых структур (спикул); миоциты — сократительные клетки; “серые клетки” — накопители гликогена. Особое место среди клеток мезоглеи занимают крупные, подвижные клетки —археошпы, ин; че — блуждающие амебоциты. Археоциты содержат в цитоплазме большое количество фагосом и лизосомальных структур, имеют большое пузырчатое ядро с хорошо различимыми ядрышками. Данные клетки очень лабильны и не имеют постоянной, строго очерченной формы. Предполагается, что этот клеточный тип тотипотентен и является предшественником всех других классов клеток у губок.
Хоаноциты, обеспечивая движение воды, одновременно фиксируют на своей поверхности органические остатки и микроорганизмы. Такой фиксированный материал захватывается затем блуждающими амебоцитами. Поглощенные микроорганизмы перевариваются , а продукты катаболизма транспортируются по всему организму.
У губок видов Terpios zeteki и Ephydatla fluviatiiis описаны два типа фагоцитирующих клеток: археоциты и колленциты. Они поглощают в условиях эксперимента широкий набор веществ: китайскую тушь, зерна кармина, эритроциты и пр.
Кишечнополостные (Coelenterata). Кишечнополостные (медузы, кораллы, морские анемоны) — двухслойные многоклеточные, обладающие истинно тканевыми образованиями в виде экто- и эндодермы. Между этими слоями находится неструктурированная, желеобразная мезоглея. Среди клеток, входящих в тканевые слои, описаны такие формы как стрекательные клетки, реснитчатые клетки, гликоциты, интерстициальные клетки. В мезоглее представлены в небольшом количестве блуждающие амебоциты, которые локализуются также в эндодерме, окружающей пищеварительный тракт. Помимо пищеварительной функции, блуждающие амебоциты принимают участие в трансплантационном отторжении. Другими клетками, претендующими на роль защитных клеточных элементов, являются подвижные интерстициальные клетки эктодермы.
Кольчатые черви (Aimelides). Кольчатые черви — животные, значительно продвинутые в филогенетическом ряду. Они обладают вторичной полостью тела (целомом) и замкнутой кровеносной системой. Целомоциты червей делят на пять основных классов: базофилы и нейтрофилы (гиалиновые амебоциты), гранулоциты и ацидофилы (гранулярные амебоциты) и хлорагогенные клетки (элеоциты). Данные электронной микроскопии позволяют отнести базофилы к лимфоцитоподобным клеткам двух типов: с большим или меньшим количеством псевдоподий (рис.20.2). Эти клетки демонстрируют структурное сходство с незрелыми лимфоцитами позвоночных и обладают маркером Т-клеток млекопитающих. Нейтрофилы напоминают макрофаги позвоночных животных (рис.
- . В их вакуолях заметны различного рода включения, в том числе и бактериальные клетки. У кольчатых червей процесс фагоцитоза, инкапсуляции, распознавания чужеродности, трансплантационного отторжения и адоптивного переноса обеспечивается в
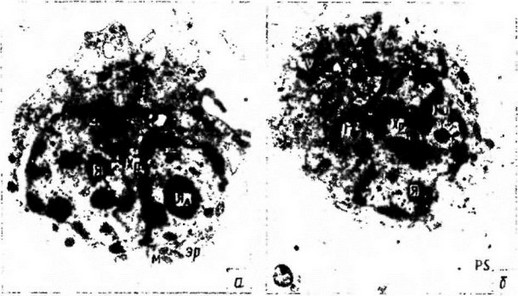
Рис. 20.2. Клетки из целомической жидкости дождевого червя Lumbricus terrestris. а — лимфоцитоподобный целомоцит типа I; б — он же, тип II. Видны: ядро (Я), плотное ядрышко (Яд), участки диффузного хроматина (Хр), шероховатый эндоплазматический ретикулум (ЭР), небольшое количество митохондрий (М), хорошо представленный аппарат Гольджи (Г), вакуоли (В), псевдоподии (ПС) (по: Cheng, 1981)
основном гиалиновыми амебоцитами: лимфоцитоподобными амебоцитами первого и второго типов (базофилами), а также нейтрофилами, характеризующимися способностью к активному фагоцитозу. При отторжении алло- и ксенотрансплантатов в реакцию вступают в основном лимфо- цитоподобные амебоциты и нейтрофил ы-макрофаги.
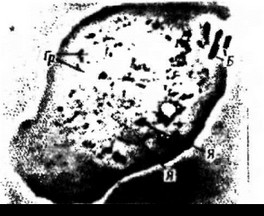 Членистоиогие (Arthro- пщодобиая клети) Lumbricus terrestrb, со- poda). ЧленИСТОНОГИе — наибо- держащий фагоцитированные бактерии (Б), лее развитый в эволюционном гРанУлы (гр)gt; (Я)» ядрышко (Яд)
Членистоиогие (Arthro- пщодобиая клети) Lumbricus terrestrb, со- poda). ЧленИСТОНОГИе — наибо- держащий фагоцитированные бактерии (Б), лее развитый в эволюционном гРанУлы (гр)gt; (Я)» ядрышко (Яд)
отношении тип первичноротых, ведущий свое начало от кольчатых червей. У хорошо изученного класса насекомых (Insecta) идентифицировано шесть типов лейкоцитов с той или иной формой иммунологической активности.
Прогемоциты напоминают малые лимфоциты позвоночных животных. Эти клетки диаметром 8-9 мкм имеют круглую или овальную форму, содержат большое ядро. Цитоплазма располагается достаточно узкой полосой вокруг ядра и лишена значительных клеточных включений и органелл. Данные клетки составляют около 1% от общего количества лейкоцитов.
Плазматоциты, составляющие 45% циркулирующих лейкоцитов, являются высокоплейоморфными клетками. Называются они разными авторами по разному: фагоциты, гранулоциты, мак- ронуклеоциты, ламеллоциты. Их ядра занимают половину объема клетки. Цитоплазма содержит гранулы различной величины, митохондрии, лизосомы, микротрубочки. Плазматоциты являются наиболее активными фагоцитирующими клетками и проявляют свойства, характерные для макрофагов позвоночных: адгезируют на стекле, активируются в присутствии чужеродных эритроцитов, выпуская псевдоподии и образуя розетки с этими эритроцитами. Кроме того, они способны реагировать на антигенные агрегаты и образуют вокруг чужеродного материала меланизированные капсулы.
Моллюски (Mollusca). Моллюски представляют собой большой, четко очерченный тип первичноротых животных. Лейкоциты моллюсков, имевшие по терминологии различных авторов до 76 названий, делятся в настоящее время на два класса: гиалиновые клетки и гранулоциты. Округлые лимфоцитоподобные клетки (рис.
- , циркулирующие в гемолимфе, способны образовывать ро
зетки с эритроцитами барана, так же как Т-клетки человека. Другая группа циркулирующих гиалиновых гемоцитов адгезиру- ет на стекле, обладает способностью к распластыванию, активно фагоцитирует инертные частицы, отвечает кислородным взрывом на стимуляцию. По этим признакам отмеченная группа клеток напоминает макрофаги позвоночных животных. Данный тип клеток наиболее активен в фагоцитозе бактерий и инкапсуляции паразитов. Подобной активностью обладают и грануло- циты, хотя и в менее выраженной форме.
 Иглокожие (Echinodermata). Иглокожие — высокоорганизованные беспозвоночные, хотя в ветви вторичноротых занимают низшее положение. В их крови и целомической жидкости представлены те же формы лейкоцитов, что и у насекомых, однако имеется и дополнительный клеточный тип — вибрирующие клетки, вооруженные жгутиками и содержащие мукополисахаридные гранулы. Эти клетки коагулируют и разрушаются в процессе изоляции чужеродного корпускулярного материала, выполняя тем самым барьерную, защитную функцию.
Иглокожие (Echinodermata). Иглокожие — высокоорганизованные беспозвоночные, хотя в ветви вторичноротых занимают низшее положение. В их крови и целомической жидкости представлены те же формы лейкоцитов, что и у насекомых, однако имеется и дополнительный клеточный тип — вибрирующие клетки, вооруженные жгутиками и содержащие мукополисахаридные гранулы. Эти клетки коагулируют и разрушаются в процессе изоляции чужеродного корпускулярного материала, выполняя тем самым барьерную, защитную функцию.
У иглокожих трансплантат инфильтруется лимфоцитами, гра- нулоцитами и фагоцитирующими мононуклеарами, что указывает на их иммунологическую активность.
Оболочники (Tunicata). Оболочники являются уникальной группой животных в силу своебразия своего роазвития. В личиночной стадии они обладают хордой, однако во взрослом состоянии в результате метаморфоза хорда полностью резорбируется. Наличие личиночной хорды ставит оболочников в единый филогенетический ряд с позвоночными животными. В связи с подобной общностью данный тип животных интересен и для понимания эволюции иммунитета как обладатель предсуществующей системы специфической защиты позвоночных.
Лимфоциты оболочников рассматриваются в качестве гомологов лимфоцитов позвоночных животных. Этот тип клеток, как и у позвоночных, чувствителен к облучению, отвечает на митогены Т-клеток, образует розетки с эритроцитами барана и обеспечивает отторжение трансплантата, инфильтруя пересаженную ткань.
Типичными клетками гемолимфы (около 70% от общего количества) являются вакуолизированные, содержащие ванадий клеточные формы. Их функция связана с переносом кислорода. Неспецифическая иммунологическая активность данных клеток проявляется в инкапсуляции инородных частиц, включая гетерологичные эритроциты. Способностью к фагоцитозу обладают многие клеточные формы: амебоциты, гиалиновые клетки, грануло- циты, макрофаги.
Позвоночные (Vertebrata). Позвоночные животные в целом характеризуются завершенностью формирования макрофагальнолимфоцитарного клеточного комплекса. В сравнении с беспозвоночными усложнение клеточного состава комплекса у позвоночных касается в основном лимфоидных клеток, но оно не связано с появлением новых морфологических типов и обогащено лишь формированием функционально различных субпопуляций в лимфоидном ряду развития.
В классе круглоротых (Cyclostomata) произошло важное эволюционное событие, которое обеспечило прогресс в становлении всего специфического иммунитета. Это — возникновение морфологических структур, явившихся прообразом тимуса более высокоорганизованных позвоночных животных. Наличие тимуса позволяет говорить о новом типе лимфоцитов — тимусзависимых клетках (Т-клетках). Кроме того, именно в этом классе впервые в филогенезе появляются морфологически оформленная костномозговая ткань протопозвоночной дуги и антителопродуцирующие плазмоциты, что указывает на возникновение на уровне круглоротых В-системы иммунитета и В-лимфоцита как самостоятельного клеточного типа.
Более чем столетний период изучения клеток ли мфо-миело- идного комплекса у рыб (Piesces) показал, что его клеточный состав представлен наиболее полно и характерен для всех вышестоящих классов позвоночных животных, включая млекопитающих. Лейкоциты крови рыб подразделяются на две основные группы: гранулоциты, включающие нейтрофилы, эозинофилы,базофилы, и агранулоциты, представленные моноцитами-макрофагами, большими, средними и малыми лимфоцитами, а также тромбоцитами.
Содержание фагоцитирующих мононуклеаров в крови рыб незначительно и колеблется от 4 до 7%. Так же, как и у других животных, макрофаги совместно с нейтрофилами обеспечивают неспецифическую иммунную защиту, а также принимают участие в воспалительных реакциях. В зоне воспаления макрофаги могут сливаться, образуя гигантские многоядерные клетки. Кроме того, фагоцитирующие мононуклеары рыб, как и млекопитающих, способны к презентации антигена, демонстрируя тем самым свое уча
стие в процессах клеточного взаимодействия при формировании специфического иммунного ответа.
Более 70% лейкоцитов периферической крови рыб представлены лимфоцитами. Популяция этих лимфоцитов функционально неоднородна и представлена как В-, так и Т-клетками. Более того, Т-клетки делятся на субпопуляции с цитотоксической, хелпер- ной и супрессорной активностью. Таким образом, лимфоидный клеточный комплекс рыб достаточно гетерогенен и фактически неотличим от такового наиболее высокоорганизованных позвоночных животных.