
При изучении любой биологической проблемы исследователь невольно обращается (или, по крайней мере, должен обращаться) к ее сравнительно-историческим аспектам. Связано это в первую очередь с тем, что изучение механизмов какого-либо явления только у млекопитающих, как правило, сталкивается с определенными трудностями в силу эволюционно сложившейся мно- гофакториал ьности процессов, лежащих в основе любого биологического феномена. Путь, который помогает изучить отдельные элементы процесса, установить их взаимосвязь и тем самым придти к пониманию явления в целом, — это обращение к филогенетически менее организованным формам жизни с постепенным переходом от групп, в которых данное явление только зарождалось, к ipynnaM, в которых оно усложнилось.
Главные вопросы эволюционной иммунологии — это решение проблемы возникновения способности к специфическому антигенному распознаванию, т.е. появления антигенраспознающих рецепторов как молекулярных факторов такого распознавания, и выяснение путей эволюционного происхождения лимфоцитов — основных участников иммунологических событий.
Кроме того, имеется более общий и фундаментальный вопрос, связанный с оценкой роли специфического иммунитета в эволюции многоклеточных животных. Как отмечали Н.В.Тимофе- ев-Ресовский и соавторы (1969), эволюционное изучение какого- либо биологического явления может внести определенный вклад в понимание, а возможно, и в изменение теоретических взглядов на течение эволюционного процесса.
Подобные представления как нельзя лучше согласуются с оценкой роли иммунитета в прогрессивном эволюционном развитии жизни. В свое время нами был выдвинут тезис о том, что на эволюцию специфического иммунитета не следует смотреть только как на самостоятельное явление исторического развития; скорее, ее следует оценивать как такой процесс, который обеспечил прогресс в мире животных по линии увеличения абсолютного количества соматических клеток.
Основной прием эволюционной иммунологии — сравнительный, использующий сопоставление конкретной формы иммунной реактивности у представителей одной определенной систематической группы с таковой у других групп, эволюционно удаленных в той или иной степени от исходной. В связи с этим филогенетическое древо — схематизированный образ возможных эволюционных преобразований в мире животных — для сравнительного иммунолога выступает в качестве матрицы, в ячейки которой закладывается информация о состоянии иммунитета определенной таксономической труппы.
На рис. 20.1 изображено устоявшееся представление о филогенетических связях в мире животных. Черными кружками отмечены те группы животных, у представителей которых анализировались определенные формы иммунной реактивности.
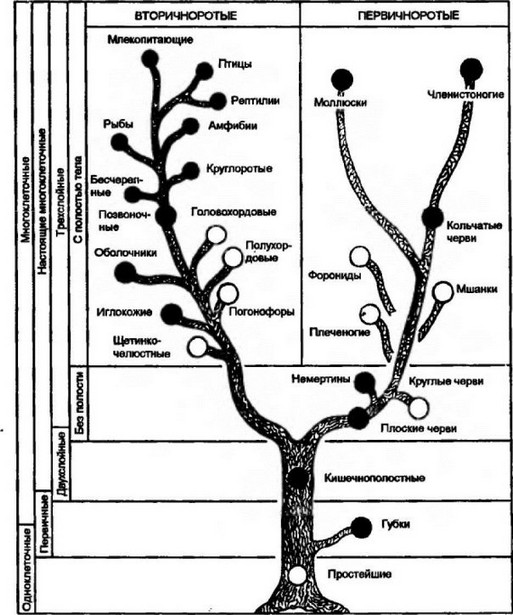
Рис. 20.1. Филогенетическое древо взаимных отношений между основными типами животных.
Черными кружками отмечены типы, у представителей которых изучались те или иные формы иммунитета.
До середины 60-х годов иммунологи проводили резкую грань между способом иммунной защиты у беспозвоночных и позвоночных животных. Считалось, что все беспозвоночные, включая хордовых, обладают только неспецифической формой реагирования, в то время как позвоночные животные, начиная с круглоротых, являются носителями приобретенного иммунитета, характеризующегося специфичностью в распознавании антигена, инду- цибельностью и формированием иммунологической памяти. Подобные различия в иммунной реактивности между двумя группами животных связывали с наличием у позвоночных тимуса, отсутствующего у беспозвоночных. Предполагалось, что для беспозвоночных с короткой продолжительностью жизни и значительным потенциалом к самовоспроизведению достаточной является неспецифическая форма реагирования. Действительно, неспецифическая клеточная и гуморальная защита у беспозвоночных выражены достаточно хорошо. При этом представления о том, что беспозвоночные лишены каких-либо проявлений специфического иммунного реагирования, оказались ошибочными.
В середине 60-х годов была показана способность представителей семейства дождевых червей Eisenia foetida и Lumbricus terrestris к специфическому отторжению аллогенной ткани и формированию кратковременной иммунологической памяти. Несколько позднее, в 70-х годах установлено специфическое аллоиммунное отторжение даже у таких низкоорганизованных многоклеточных, каковыми являются губки и кишечнополостные. Полученные факты ясно указывали на то обстоятельство, что специфический иммунитет не является прерогативой только позвоночных животных. Первые иреадаптационные проявления специфического иммунного реагирования возникли очень давно, вероятно, с момента появления первых многоклеточных, около 2 млрд, лет тому назад. Кроме того, и на это следует обратить особое внимание, выявленные факты позволяют предполагать время возникновения предшественников антигенраспознающих рецепторов Т- и В-клеток.
Особое место в разработке проблем эволюции иммунитета занимает вопрос о происхождении лимфоцита — основного участника иммунологических событий. Вполне вероятно, что лимфоцит как самостоятельный клеточный тип исторически возник специально для осуществления клеточной формы иммунного реагирования, а понятия “лимфоидный” и “иммунный” — синонимы для обозначения одной и той же системы организма. Подобная оценка строится на факте одновременного появления в эволюции нового клеточного типа — лимфоцита — у немертин и кольчатых червей и способности этих животных к специфическому отторжению трансплантата.
Морфологический аспект эволюции лимфоидной системы представляет самостоятельную проблему, в равной мере относящуюся как к гематологии, так и иммунолгии. По представлениям В.Н.Беклемишева (1964), родоначальным клеточным типом, давшим начало внутренней (мезенхимальной) среде организма, был блуждающий амебоцит кишечнополостных. Факт появления амебоцита-макрофага автор оценивал как важное арогенное событие, следствием которого явилось формирование целой системы органов, объединенных в лимфо-миелоидный комплекс. Элементы комплекса в виде различного рода лимфоидных скоплений и узелков уже представлены у беспозвоночных с полостью тела: кольчатых червей, моллюсков, членистоногих, иглокожих, оболочников. Однако своего совершенства комплекс достигает у высших позвоночных животных.
Обращаясь собственно к лимфоидной системе, следует выделить, по крайней мере, два основных исторических события, два эволюционных новшества:
- возникновение лимфоцита как морфологически обособленного клеточного типа, основная функция которого — иммунологическая;
- вычленение из лимфо-миелоидного комплекса лимфоидной ткани в качестве автономной структуры со своими специфическими (иммунологическими) функциями; особое место здесь занимает тимус.