
Димеры ap и у5 — собственно антигенраспознающая часть TCR (см. табл. 3.2). Они имеют аналогичное строение и молекулярную организацию. Полипептидные цепи, входящие в состав TCR, принадлежат к суперсемейству иммуноглобулинов. Димеры TCR организованы значительно проще, чем иммуноглобулины. Цепи TCR по строению сходны с L-цепями иммуноглобулинов. Каждая цепь TCR имеет 2 внеклеточных домена, один из которых вариабельный, а другой (расположенный ближе к мембране) — константный (рис. 3.8). Домены содержат 87—113 аминокислотных остатков. Гомология полипептидных цепей TCR и иммуноглобулинов невелика, но они имеют сходную доменную организацию. Домены цепей TCR образованы двумя p-слоями. В константном домене они содержат 4 и 3, а в вариабельном — по 4 p-складки. Во всех типах полипептидных цепей TCR в каждом из двух доменов содержится по одному участку гликозилирования. В отличие от L-цепей, полипептидные цепи TCR являются мембранными молекулами, имеющими трансмембранный (10—12 аминокислотных остатков) и короткий цитоплазматический (3—5 аминокислотных остатков) участки. Цепи скреплены дисульфидной связью, расположенной непосредственно над клеточной мембраной в шарнирном участке молекул. Трансмембранные отделы цепей связаны друг с другом за счет противоположных электричес-
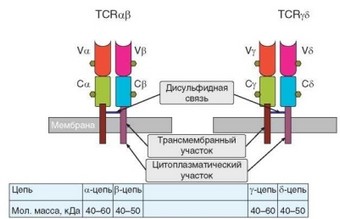
Рис. 3.8. Строение антигенраспознающих димеров Т-клеточных рецепторов
ких зарядов. Цепи димера (ар или у8) связаны с полипептидными цепями комплекса CD3, также нековалентно, на основе электростатических взаимодействий (при участии лизина в TCR и аспарагиновой кислоты в CD3). Характеристика полипептидных цепей TCR представлена в табл. 3.3.
В V-доменах TCR, как и в V-доменах иммуноглобулинов, есть 4 каркасных участка с относительно постоянной аминокислотной последовательностью и 3 — CDR. Особенно высокая вариабельность свойственна CDR3. Гипервариабельные участки двух цепей формируют антигенсвязывающую полость TCR. Как и в иммуноглобулинах, выделяют несколько семейств V-доменов TCR, различающихся строением каркасных последовательностей.
Таблица 3.3. Характеристика полипептидных цепей комплекса T-клеточный рецептор CD3
|
Цепь |
Молекулярная масса, кДа |
Число остатков |
Число цепей в комплексе |
Число внеклеточных доменов |
Число ITAM |
Число остатков Cys-Cys |
|
TCRa |
40-60 |
250-270 |
1 |
2 (SFIg — V и C) |
0 |
5-6 |
|
TCRp |
40-50 |
290-315 |
1 |
2 (SFIg — V и C) |
0 |
6 |
|
TCRy |
45-60 |
300-315 |
1 |
2 (SFIg — V и C) |
0 |
8 |
|
TCRS |
40-60 |
400-425 |
1 |
2 (SFIg — V и C) |
0 |
6 |
|
CD3y |
25 |
160 |
1 |
1 (SFIg — C) |
1 |
3 |
|
CD3S |
20 |
150 |
1 |
1 (SFIg — C) |
1 |
4 |
|
CD3e |
20 |
164 |
2 |
1 (SFIg — C) |
1 |
2 |
|
CD3Z |
16 |
142 |
1-2 |
1 |
3 |
1 |
|
CD3n |
22 |
152 |
0-1 |
1 |
3 |
1 |
SFIg — суперсемейство иммуноглобулинов.
В отличие от антител, распознающих эпитопы свободных антигенов, apTCR распознает пептидные фрагменты антигенов, встроенные в молекулы MHC. При этом антигенный пептид взаимодействует с наиболее вариабельным CDR3, а прилежащие участки молекулы MHC — с CDR1 и CDR2 (особенности распознавания антигенов разными типами рецепторов будут подробно рассмотрены далее, см. раздел 3.2.2.3). Для распознавания необходимо сохранение димерной структуры рецепторов, тогда как изолированные пептидные цепи не способны распознавать антиген. р-Цепи константных доменов полипептидных цепей TCR, относящиеся к различным семействам, различаются по способности связывать суперантигены (см. раздел 3.2.2.4). Растворимые формы TCR не образуются. Растворимые молекулы TCR, созданные генно-инженерными методами, не способны распознавать антигены. Антигенсвязывающие участки TCR обладают идио- типической специфичностью, иногда сходной с таковой антител.
В норме на поверхности Т-клетки содержится 30 000—40 000 молекул apTCR, т.е. в 4—5 раз меньше, чем BCR на поверхности В-клетки. TCR, как и CD3, являются абсолютными маркерами Т-лимфоцитов: они экспрессируются только на зрелых Т-клетках. Каждая Т-клетка может нести на поверхности только один тип TCR — ар или у8. На этой основе выделяют две главные разновидности Т-клеток, кардинально различающиеся по своим
свойствам. Большинство (gt;95%) Т-клеток, локализующихся во вторичных лимфоидных органах, циркулирующих в крови и лимфе, имеют рецептор ав-типа. у5Т-клетки составляют минорную субпопуляцию (1—3%), представленную преимущественно в барьерных тканях, особенно в слизистой оболочке кишечника, где их содержание достигает 20—30% от общего числа Т-лимфоцитов.