
В процесс распознавания Т-клетками антигена (точнее, комплекса антигенного пептида с молекулой MHC), наряду с антигенраспознаю- щим рецепторным комплексом вовлекаются дополнительные молекулы. Наиболее важные из них — корецепторы CD4 и CD8. Назначение этих молекул состоит прежде всего в повышении сродства рецепторного комплекса к лиганду за счет дополнительного связывания корецепторов с молекулами MHC (отсюда обозначение молекул как корецепторов). Поскольку антигенный пептид может презентироваться Т-клетке в составе молекул MHC, относящихся к двум разным классам — I и II, в распознавании могут участвовать две разновидности корецепторов — CD8 и CD4. Молекула CD8 обладает сродством к MHC-I, а молекула CD4 — к MHC-II. Поскольку на зрелых Т-клетках экспрессирован либо CD4, либо CD8, антигенные пептиды в составе молекул MHC-I распознаются CD8 + Т-клетками, а антигенные пептиды в составе молекул MHC-II — CD4+ Т-клетками. Связывание корецепторов повышает сродство TCR к антигенному комплексу в 100 раз.
СD4 и CD8 — маркеры двух главных субпопуляций авТ-клеток — соответственно, Т-хелперов и цитотоксических Т-лимфоцитов. Однако эти молекулы экспрессируются не только на клетках названных субпопуляций: CD4 в небольшом количестве выявляют на дендритных клетках и макрофагах, а CD8 — на естественных киллерах. На незрелых Т-клетках — кортикальных тимоцитах — оба корецептора экспрессируются одновременно (что определяет мембранный фенотип CD4+ CD8+). В процессе дифференциров- ки происходит супрессия гена одного из корецепторов, тогда как второй продолжает экспрессироваться.
Структурно 2 типа корецепторов существенно отличаются друг от друга, хотя оба относятся к суперсемейству иммуноглобулинов. Их свойства отражены в табл. 3.4, а схемы строения представлены на рис. 3.11. Обе молекулы состоят из трансмембранных полипептидных цепей I типа, причем СD4 — мономер, а CD8 — димер.
Молекулярная масса CD4 — 56 кДа. Внеклеточная часть единственной цепи CD4 содержит 4 иммуноглобулиноподобных домена, из которых 2 построены по типу V-, а 2 других — по типу С-доменов иммуноглобулинов. Наружный домен содержит участок, отвечающий за взаимодействие с молекулами MHC-II. В этом же домене расположен рецепторный сайт для вируса ВИЧ-1 (остатки 31—57 и 81—94 домена V1). CD4 — основной рецептор для ВИЧ, поэтому клетками-мишенями этого вируса служат CD4+ Т-лимфоциты — Т-хелперы, а также макрофаги и дендритные клетки.
Таблица 3.4. Характеристика корецепторов Т-клеток
|
Свойство |
CD4 |
CD8 |
|
Полипептидные цепи |
1 |
2 (ар). Встречается также гомодимер аа |
|
Молекулярная масса, кДа |
56 |
ар — 69 (32+37); аа — 64 (32+32) |
|
Внеклеточные домены |
4 (суперсемейство иммуноглобулинов) — V—C—V—C |
По одному (суперсемейство иммуноглобулинов) — V |
|
Сродство к молекулам MHC |
MHC-II (домен Р2) |
MHC-I (домен а3) |
|
Экспрессия на клетках |
Т-хелперы, регуляторные Т-клетки, часть NKT- и у8Т-клеток; слабо — макрофаги, дендритные клетки |
Цитотоксические Т-лимфоциты, часть у8Т-клеток и естественных киллеров |
|
Функция |
Повышение сродства к антигену-лиганду, формирование синапса, передача сигнала через Lck |
Повышение сродства к антигену-лиганду, формирование синапса, передача сигнала через Lck |
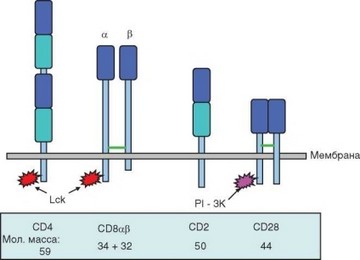
Рис. 3.11. Вспомогательные молекулы Т-лимфоцитов. Синим отмечены V-подобные домены, голубыми — С-домены; зеленые линии — дисульфидные связи, звездчатые фигуры — тирозинкиназы
Молекула CD8 — димер. На большинстве арТ-клеток CD8 — гетеродимер, образованный а- и p-цепями, на некоторых Т-клетках, локализованных преимущественно в слизистых оболочках (у человека это исключительно у5Т-клетки) — гомодимер аа. Значение существования двух типов гетеродимеров CD8 не установлено. Обе цепи CD8 сходны по размеру и строению. Их молекулярная масса составляет 34 кДа (а) и 32 кДа (Р). Во внеклеточной части цепи гомодимера образуют единственный иммуноглобулиноподобный домен, отделенный от мембраны длинным спейсерным участком. Цепи связаны нековалентно. С молекулой MHC-I взаимодействует а-цепь.
Цитоплазматическая часть и CD4 и CD8 связана с тирозинкиназой Lck (семейство Src). Прочность связи увеличивается при распознавания комплекса антигенный пептид—молекула MHC в результате конформационных изменений корецептора. Киназа Lck играет важнейшую роль в передаче активационного сигнала от TCR (см. раздел 3.5.2.1). Таким образом, CD4 и CD8 участвуют не только в распознавании комплекса антигенного пептида и молекулы MHC, но и выполняют сигнальную функцию.
Еще одна функция корецепторов реализуется в процессе презентации антигена Т-лимфоцитам. Корецепторы вместе с молекулой Lck, в отличие от комплекса TCR-CD3, входят в состав рафтов — структурно-функциональных элементов мембраны, важных для формирования иммунного синапса — временной структуры, обеспечивающей эффективное распознавание антигена Т-клетками и формирование полноценного активационного сигнала. Уже на ранних стадиях активации Т-клеток в процессе презентации антигена между комплексом TCR—CD3 и корецепторами происходит нековалентное взаимодействие, что позволяет корецептору «ввести» рецепторный комплекс в состав рафта, тем самым обусловливая эффективность презентации (см. раздел 3.5.1.3.).
Наряду с проявлением усиливающих эффектов, корецепторы способны передавать супрессорные сигналы при распознавании антигена и последующей активации клеток. Супрессорный эффект проявляется, например, при изолированном перекрестном «сшивании» молекул корецепторов моноклональными антителами. На фоне такого связывания стимуляция через TCR вызывает апоптоз Т-лимфоцитов.