
Как уже отмечено, у взрослых млекопитающих, включая человека, мие- лопоэз происходит в костном мозгу. Все клетки крови происходят от гемопоэтических стволовых клеток. Стволовые кроветворные клетки проходят 3 стадии развития, различающиеся по способности восстанавливать кроветворение при переносе их облученным животным:
- стволовые клетки длительного действия;
- стволовые клетки короткого действия;
- мультипотентные родоначальные клетки (рис. 2.1).
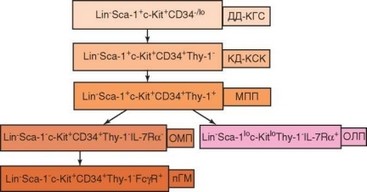
Рис. 2.1. Длительность пребывания основных миелоидных клеток в костном мозгу, крови и тканях
Для всех стадий характерен мембранный фенотип Lin-Sca-1+c-Kit+ [Lin — линейные маркеры; Sca-1 — антиген стволовых клеток (Stem cell antigen); c-Kit — лиганд фактора стволовых клеток SCF (Stem cell factor)]. Стволовые клетки на 2-й и 3-й стадии развития несут на поверхности молекулу CD34. Этот маркер чаще всего используют в качестве идентификационного для выявления стволовых клеток и их ближайших потомков. Для гемопоэтических стволовых клеток человека характерно отсутствие линейных маркеров, наличие молекулы CD34 и отсутствие молекулы CD38. Последняя появляется на стадии коммитированных предковых клеток — общих лимфоидного (CLP) и миелоидного предшественников. В костном мозгу человека содержится 0,5—5% CD34+ клеток, только 1—10% из них лишены CD38, т.е. могут рассматриваться как кандидаты в стволовые клетки. Истинных стволовых клеток, т.е. клеток, способных длительно поддерживать гемопоэз in vitro или при пересадке в организм, значительно меньше — 1 на 104 Lin-CD34+ Thy-Гклеток.
Стволовые клетки делятся медленно (около 5% находятся в S и G2 фазах клеточного цикла; каждая клетка делится 1 раз в 30—60 сут). Жизнеспособность стволовых клеток обеспечивается стромальными клетками костного мозга, формирующими их нишу. Большинство стволовых кроветворных клеток расположено в эндостальной части костного мозга, а также в синусоидах (соответственно — эндостальная и сосудистая ниши). Таких клеток мало в центральной части костного мозга. Основную роль в поддержании жизнеспособности и обеспечении функционирования стволовых клеток играют их контакты со специализированными остеобластами ниш. В зонах контакта происходит взаимодействие многих пар молекул, включая молекулы адгезии (интегрины и их рецепторы), мембранные цитокины (SCF и его лиганд, c-Kit), хемотаксические молекулы (хемокины и их рецепторы) и т.д. Роль растворимых факторов (цитокинов) в регуляции жизнеспособности и активности стволовых клеток невелика. Стволовые клетки входят в клеточный цикл и дифференцируются только при ослаблении связи с нишей и утрате контактов с остеобластами. Важная особенность стволовых клеток — сбалансированность процессов пролиферации и дифференцировки: на уровне популяции одна из дочерних клеток продолжает делиться, тогда как другая подвергается дифференцировке, то есть созревает. В результате пролиферирующие клетки образуют как бы ствол, от которого постоянно отделяются клетки, уходящие в дифференцировку.
Дифференцируясь, стволовые кроветворные клетки дают начало двум основным ветвям клеток крови — миелоидной и лимфоидной. Несколько схематизируя, можно сказать, что эти ветви обеспечивают развитие клеток соответственно врожденного и адаптивного иммунитета. В этой главе рассмотрим только миелоидный путь развития клеток крови, проиллюстрированный на рис. 2.2. Исходные клетки этого пути развития — общие миелоидные предшественники. Они образуются как в эндостальной, так и в сосудистой нишах и отличаются от стволовых клеток отсутствием мембранной молекулы Sca-1, а от CLP — рецептора для IL-7.
Из общего миелоидного предшественника происходят все клетки крови, кроме лимфоцитов. Рассмотрим только развитие клеток врожденного иммунитета по 3 линиям дифференцировки. Наиболее важная из них — грануло- цитарно-макрофагальная, или GM-линия. Она дает начало двум дочерним линиям — моноцитарной (М-линия) и гранулоцитарной (G-линия). Вопрос о развитии эозинофилов и базофилов до конца не решен. Есть данные о наличии у них общего предшественника, дифференцирующегося впоследствии на эозинофильную и базофильную линии. С помощью традиционного морфологического подхода выделяют несколько стадий развития миело- идных клеток: миелобласты, промиелоциты, миелоциты, метамиелоциты, палочкоядерные и сегментоядерные (зрелые) формы. Клетки на 2 последних стадиях в норме присутствуют в кровотоке. Накапливается все больше свидетельств, что традиционное представление гемопоэза в виде «дерева» не точно, поскольку описаны многочисленные отклонения от сложившейся схемы. Так, известны клетки-предшественники, общие для моноцитов и В-лимфоцитов или моноцитов и Т-лимфоцитов. Гемопоэзу свойствена значительная степень пластичности.
Для дифференцировки миелоидных (как и любых других) клеток необходима экспрессия определенного набора факторов транскрипции. Эти ядерные белки обладают сродством к конкретным последовательностям ДНК в промоторных участках генов и, соединяясь с ними, обеспечивают экспрессию этих генов. Обычно с промотором соединяется целый комплекс транскрипционных факторов, среди которых есть постоянно присутствующие (конститутивные) и индуцируемые факторы; некоторые из них характерны для различных стадий развития клеток. Так, при миелопоэзе для большинства линий и стадий развития клеток выявлены относительно специфичные для них транскрипционные факторы (см. рис. 2.2). Например, экспрессия транскрипционного фактора Ikaros характерна для лимфоидной, но не миелоидной ветви гемопоэза. При миелопоэзе высокий уровень экспрессии фактора PU.1 необходим для развития клеток GM-линии, низкий уровень выявляют в клетках эозинофильного ряда, а в базофилах этот фактор отсутствует. Моноцитарный и гранулоцитарный ряды различаются скорее комбинацией транскрипционных факторов, чем наличием одного специфичного; нейтрофилы отличаются от моноцитов экспрессией разных
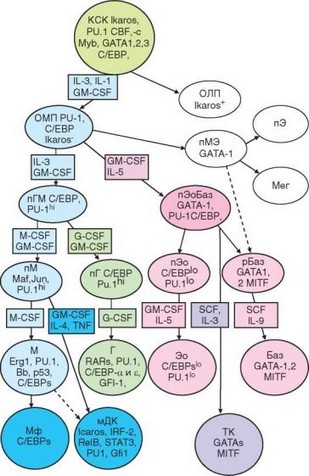
Рис. 2.2. Кроветворные стволовые клетки мышей и ранние этапы их дифференци- ровки. В прямоугольниках отмечены цитокины, ответственные за развитие клеток, в овалах — обозначения клеток и специфичных для них транскрипционных факторов. Обозначения клеток см. в тексте
изоформ фактора C/EBP. Спектры дифференцировочных факторов базофилов и других миелоидных клеток крови не перекрываются.
Незрелые кроветворные клетки легко подвергаются апоптозу. Для сохранения жизнеспособности им необходимо присутствие в микроокружении цитокинов. Основным цитокином, общим практически для всех миелоидных клеток, начиная от общего миелоидного предшественника, считают гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF). На ранних этапах миелопоэза сходную роль выполняет IL-3, называемый также полипоэтином. При созревании и специализации клеток для сохранения жизнеспособности им необходимы линейно-специфические цитокины: для моноцитарного ряда — моноцитарно-макрофагальный колониестимулирующий фактор (M-CSF), а для нейтрофильного — грану- лоцитарный колониестимулирующий фактор (G-CSF). Подобную роль при развитии эозинофилов играет IL-5. Во всех этих случаях наряду с названными цитокинами роль фактора выживания и колониестимулирующего фактора выполняют GM-CSF и, в меньшей степени, IL-3. Базофилам для развития нужен комплекс факторов, в котором главную роль играет CSF.
Между появлением гранулоцитарно-макрофагального предшественника и его дифференцировкой на моноцитарно-макрофагальный и гранулоци- тарный предшественники проходит около 5 сут — это один из самых длительных этапов миелопоэза. Следующий этап — созревание — значительно отличается по продолжительности для моноцитов и гранулоцитов: если для созревания моноцита необходимо 2—3 сут, то для созревания нейтрофильного гранулоцита — 10—12 сут. После созревания моноциты находятся в костном мозгу еще сутки и затем покидают его, поступая в кровоток. При этом клетки сохраняют способность к делению и дальнейшей дифференци- ровке. Нейтрофилы остаются в костном мозгу в течение 1—2 сут и выходят в кровь не просто зрелой, а старой клеткой с ограниченными способностями, неспособной к делению, индуцированной экспрессии генов и синтезу белка. Наиболее короткий промежуток времени требуется для развития в костном мозгу эозинофилов (2—4 сут). Аналогичные данные для базофилов отсутствуют. Продолжительность пребывания различных миелоидных клеток в костном мозгу, кровотоке и тканях сопоставлена на рис. 2.3.
Выход лейкоцитов из костного мозга в кровоток происходит вследствие ослабления взаимодействия хемокинов, выделяемых стромальными клетками костного мозга с рецепторами лейкоцитов. Наиболее важный хемокин, удерживающий созревающие клетки в костном мозгу, — CXC12 (SDF-1 — Stroma derived factor 1, фактор стромальных клеток 1), распознаваемый рецептором CXCR4 (о хемокинах и их рецепторах см. раздел 2.3.2). Под влиянием колониестимулирующих факторов (гемопоэтинов) происходит ослабление выработки хемокинов и экспрессии их рецепторов, что позволяет созревшим клеткам покинуть костный мозг. Сегментоядерные (нейтрофильные и эозинофильные) лейкоциты пребывают в кровотоке менее 12 ч; моноциты
|
Клетки |
Развитие в КМ |
Циркуляция в крови |
Пребывание в тканях |
|
Моноциты-Мф |
8-9 сут |
1-2 сут |
20-25 сут - месяцы/годы |
|
Нейтрофилы |
18-20 сут |
7-10 ч |
3-5 сут |
|
Эозинофилы |
8-10 сут |
5-10 ч |
10-12 сут |
Рис. 2.3. Схема миелопоэза с указанием цитокинов, ответственных за выживание и развитие клеток и дифференцировочных факторов
циркулируют в течение нескольких дней. Затем клетки мигрируют из крови в ткани. Этот процесс регулируется хемокиновыми сигналами и происходит с участием молекул адгезии (селектинов, интегринов) и их рецепторов. В норме экстравазация лейкоцитов осуществляется по тем же законам, что и при воспалении, но менее интенсивно в связи с меньшей проницаемостью сосудистой стенки и более слабой хемокиновой стимуляцией.
Длительность пребывания миелоидных клеток в тканях также существенно варьирует: для нейтрофилов и эозинофилов она значительно меньше, чем для моноцитов. Так, в тканях нейтрофилы живут всего 3—5 сут, эозинофилы — 10—12 сут, тогда как моноциты (точнее, макрофаги, в которые они превращаются в тканевом микроокружении) могут находиться в тканях до нескольких лет (для разных субпопуляций макрофагов этот показатель существенно различается). Нейтрофилы, эозинофилы и базофилы мобилизуются из крови в ткани в особых экстренных случаях (острое воспаление, аллергические процессы). Моноциты/макрофаги, наоборот, играют преимущественно роль клеток, длительное время живущих и функционирующих в различных тканях. В связи с этим нужно отметить значительно более высокую производительность гранулоцитопоэза по сравнению с моноцито- поэзом. За сутки в организме человека образуется и поступает в кровоток около 1011 нейтрофилов, что по массе составляет около 100 г, т.е. примерно 0,1% от массы тела; такое же количество гранулоцитов ежедневно погибает. Производительность моноцитопоэза в 20 раз ниже: за сутки образуется и поступает в кровоток до 5х109 моноцитов. Это обусловлено значительно большей продолжительностью жизни моноцитов/макрофагов.
Существует еще по крайней мере 2 разновидности миелоидных клеток, происходящих из костного мозга, но использующих кровяное русло только в качестве кратковременного транзитного участка по пути в ткани — дендритные и тучные клетки. Развитие этих клеток будет рассмотрено отдельно при описании их функций (см. разделы 2.1.4 и 2.1.6).