
Моноциты и макрофаги представляют стадии развития миелоидных клеток. Они образуют мононуклеарную фагоцитирующую систему. Роль макрофагов в качестве одних из основных фагоцитирующих клеток установлена в 1882 г. И.И. Мечниковым, давшим этим клеткам их название. В 20-е гг. ХХ века Л. Ашоф (L. Aschoff) создал учение о ретикуло-эндотели- альной системе — защитной системе, объединяющей тканевые фагоцитирующие клетки. Позже для обозначения практически той же системы стали использовать термин «мононуклеарная фагоцитирующая система».
Циркулирующий вариант клеток — моноцит, тканевый — макрофаг. Превращение моноцита в макрофаг происходит под влиянием тканевого микроокружения и сопровождается экспрессией новых генов, т.е. может рассматриваться как дифференцировка клеток. Эту дифференцировку регулирует M-CSF. Моноциты представляют довольно крупные клетки (диаметром 9—15 мкм) с ядром бобовидной формы и тонкой структурой хроматина. Макрофаги значительно крупнее моноцитов (диаметр составляет 20—25 мкм) и имеют распластанную форму. В отличие от округлых моноцитов, макрофаги имеют неправильные очертания и морфологически полиморфны.
Часто морфологических признаков оказывается недостаточно для дифференциации моноцитов от макрофагов, а также для определения их разновидностей. В этом случае используют дополнительные приемы, такие как определение ферментов или мембранных молекул-маркеров (рис. 2.5, 2.6). Наиболее важные в функциональном отношении мембранные молекулы моноцитов/макрофагов — рецепторы, предназначенные для распознавания PAMP (см. подробнее раздел 2.2), в первую очередь — толл-подобные рецепторы (TLR) (см. табл. 2.1). Ни на каких других клетках эти рецепторы не представлены так разнообразно, как на моноцитах и макрофагах. Это обеспечивает макрофагам и моноцитам возможность распознавать фактически все основные группы паттернов. На этих клетках обнаружены все разновидности TLR, для которых характерна экспрессия на поверхности клеток — TLR-1, TLR-2, TLR-4, TLR-5, TLR-6, TLR-11. С мембранным рецептором TLR-4 функционально связан один из основных маркеров моноцитов и макрофагов — молекула CD14. CD14 взаимодействует с комплексом бактериального ЛПС с ЛПС-связывающим белком, что облегчает взаимодействие Л ПС с TLR-4. TLR, распознающие чужеродные нуклеиновые кислоты (TLR-3, TLR-7, TLR-8, TLR-9), локализованы внутриклеточно — на мембранах цитоплазматических гранул. К группе мембранных молекул, распознающих паттерны, следует отнести молекулу CD13 (аминопептидаза N), характерную для моноцитов, но не макрофагов. Как уже было отмечено, CD13 обладает сродством к антигенам оболочки ряда вирусов.
Для моноцитов/макрофагов свойственна также экспрессия других рецепторов врожденного иммунитета — лектиновых. Лектиновые рецепторы моноцитов и макрофагов распознают свободные D-гликозильные остатки глюкозы, галактозы, маннозы, фукозы, N-ацетилглюкозамина, N-ацетилгалактозамина. На собственных клетках организма такие остатки экранированы остатками сиаловой кислоты и экспонируются преимущественно на старых клетках, подлежащих элиминации. Патогены
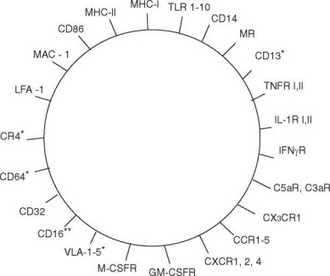
Рис. 2.5. Основные мембранные молекулы макрофагов. * В основном моноциты. ** В основном макрофаги
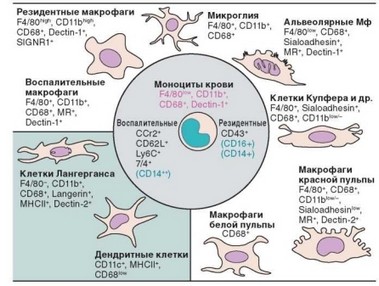
Рис. 2.6. Фенотипическая гетерогенность моноцитов/макрофагов и дендритных клеток как результат дифференцировки in vivo. Внутри круга — моноциты и их разновидности, вне круга — разновидности макрофагов. Указаны мембранные маркеры клеток
(бактерии, грибы, простейшие) несут на поверхности гликоконъюгаты с неэкранированными гликозильными остатками, распознаваемыми лек- тиновыми рецепторами макрофагов, что облегчает фагоцитоз патогенов. В результате распознавания происходит эндоцитоз (пиноцитоз, фагоцитоз) образующихся комплексов. Наиболее важный рецептор лектиновой группы — маннозный рецептор (MR, CD206), характерный для макрофагов и слабее эксперссированный на моноцитах. И на моноцитах, и на макрофагах присутствуют лектиновые рецепторы DC-SIGN (CD209) и дектин-1. Экспрессия дектина-1 подавляется при активации макрофагов. Сигналом к фагоцитозу является также связывание с лигандами так называемых scavenger-рецепторов («мусорщиков»), к которым относят молекулу MSR (Macrophage scavenger receptor, СD36), обладающую сродством к коллагену.
Другая группа рецепторов, разнообразно представленных на моноци- тах/макрофагах, — Fc-рецепторы (молекулы, распознающие Fc-участок молекул иммуноглобулинов, обычно в связанном с антигеном состоянии). Эти рецепторы обеспечивают распознавание и облегчают фагоцитоз и разрушение моноцитами и макрофагами опсонизированных антителами клеток (в том числе патогенных); параллельно происходит активация фагоцитов. Моноциты экспрессируют полный набор Fcy-рецепторов — FcyRI (CD64), FcyRII (CD32) и FcyRIII (CD16). На макрофагах присутствуют только FcyRII и FcyRIII. Моноциты — единственный тип клеток, спонтанно экспрессирующих FcyRI, обладающий наиболее высоким сродством к молекуле IgG и способный связывать его даже в свободном состоянии, а не только в составе иммунного комплекса, как это происходит обычно. На моноцитах и макрофагах представлены также рецепторы для Fc-части IgA (FcaR) и низкоаффинные рецепторы для IgE — FceRII (CD23). Эти рецепторы участвуют в регуляции синтеза антител соответствующих изотипов.
Благодаря присутствию на поверхности моноцитов и макрофагов рецепторов для комплемента (CR) эти клетки распознают фрагменты факторов комплемента, прикрепленные к поверхности патогенов. Большинство рецепторов распознает фрагменты C3b и C3d — CR1 (CD35), CR3 (CD11b/ CD18, или Мас-1) и CR4 (CD11c/CD18, или р150,95). Функция этих рецепторов сходна с таковой Fc-рецепторов: они облегчают распознавание клеток- мишеней фагоцитами и поставляют в фагоцитирующие клетки активационные сигналы. Моноциты/макрофаги экспрессируют также рецепторы для фактора C1q и хемотаксических факторов-анафилатоксинов C3a и C5a.
Поскольку для проявления функциональной активности моноцитам/ макрофагам важно взаимодействие с межклеточным матриксом (в процессе миграции) и с другими клетками (при участии в реакциях иммунитета), на их поверхности представлено большое число молекул адгезии. Среди них особенно важны интегрины (см. раздел 2.3.1.2), например ргинтегрины, обеспечивающие связи с молекулами межклеточного матрикса (коллагеном, фибронектином, ламинином). Из ргинтегринов на моноцитах экспрессированы VLA-4, VLA-2, VLA-5 и VLA-6; 3 последних на макрофагах отсутствуют. VLA-2, VLA-5 и VLA-6 взаимодействуют с названными молекулами матрикса, а VLA-4 — еще и с мембранной молекулой лимфоцитов и самих активированных макрофагов VCAM-1. Все 3 р2-интегрина — LFA-1, Mac-1 (CR3) и p150,95 (CR4) — присутствуют на поверхности как моноцитов, так и макрофагов. Р2-Интегрины взаимодействуют преимущественно с интег- риновыми рецепторами ICAM-1 (CD54), ICAM-2 (CD102) и ICAM-3 (CD50), присутствующими как на Т-лимфоцитах, так и на самих макрофагах (особенно после их активации), а также на активированных эндотелиальных и эпителиальных клетках. За адгезию к эндотелиальным клеткам, необходимую при транссосудистой миграции, отвечает молекула PЕCAM (CD31). К молекулам адгезии необходимо отнести также CD15 (Lewis X) — углеводный компонент мембранных гликоконъюгатов (разветвленный трисахарид), служащий рецептором для молекул адгезии селектинов, который распознает углеводы (см. раздел 2.3.1.1).
Функционально важную группу поверхностных молекул моноцитов/ макрфоагов образуют молекулы MHC (см. раздел 3.2.2.2) и костимулирую- щие молекулы (см. раздел 3.4.1.4). Роль МНС состоит в представлении (презентации) антигенных пептидов TCR. Если молекулы МНС-I присутствуют на всех ядросодержащих клетках организма, то молекулы МНС-II экспрессированы только на специализированных АПК, к которым наряду с дендритными клетками и В-лимфоцитами относят макрофаги. Экспрессия молекул МНС-II усиливается при активации клеток. Презентация антигена — узловое событие иммунного ответа, связывающее реакции врожденного и адаптивного иммунитета. В ходе презентации молекула МНС распознается как самим TCR, так и корецепторами — СD8 и СD4, обладающими сродством к молекулам МНС-I и MHC-II соответственно. Молекула CD4 в небольшом количестве экспрессирована на некоторых макрофагах, что делает их чувствительными к инфицированию вирусом иммунодефицита человека (ВИЧ), для которого молекула CD4 служит основным рецептором (см. раздел 4.7.2). Помимо презентации антигена, для эффективной активации Т-клеток необходима их костимуляция. Она достигается при взаимодействии пар молекул АПК и Т-лимфоцита, называемых костиму- лирующими. Со стороны АПК (в том числе макрофага) в роли костиму- лирующих выступают молекулы CD80 и CD86. Первая из них появляется на поверхности клетки только после активации, вторая экспрессируется конститутивно (даже на покоящихся клетках), но при получении активационного сигнала ее экспрессия усиливается.
Важная группа мембранных молекул моноцитов/макрофагов — рецепторы для цитокинов. Из них наиболее специфичен для моноцитов и макрофагов Fms (CD115) — рецептор для их линейного фактора M-CSF. Наличие Fms позволяет дифференцировать моноциты и их предшественники от клеток гранулоцитарного ряда, на которых этот рецептор отсутствует. Для проявления макрофагами их функций, как эффекторных клеток иммунитета, особенно важны рецепторы для интерферона у (IFNyRI и IFNyRII — CD119); для провоспалительных цитокинов (которые они сами же и секре- тируют) IL-1 (CD121a, CD121b) и TNF (CD120a, CD120b); а также рецепторы для IL-6, IL-12, IL-18, колониестимулирующего фактора GM-CSF (CD116) и ряда других цитокинов. Высокая подвижность моноцитов и особенно макрофагов требует экспрессии рецепторов для хемотаксических факторов. Некоторые из них (рецепторы для С3а и С5а) уже упоминались. Моноциты и макрофаги располагают широким спектром рецепторов для специали-
зированных хемотаксических цитокинов — хемокинов (см. раздел 2.3.2.2), особенно провоспалительных: CXCR1, ^R1, CCR2, CCR3, CCR4, CCR5, CCR8, CX3CR1.
При гистохимической идентификации моноцитов/макрофагов определяют наличие в клетках неспецифической эстеразы, диффузно распределяющейся в цитоплазме. Выявление ряда ферментов позволяет оценить степень зрелости этих клеток. Так, например, миелоперокидаза содержится в значительном количестве в моноцитах; их превращение в макрофаги сопровождается утратой этого фермента. Экспрессия 5’-нуклеотидазы, Р-галактозидазы и аминопептидазы при этом, наоборот, возрастает, а трансглутаминазу удается выявить только в зрелых макрофагах. Помимо названных, в макрофагах присутствуют и другие ферменты — коллагена- за, протеиназы, липазы, нуклеазы, фосфатазы и др. Некоторые ферменты макрофагов участвуют в реализации бактерицидной активности: кислород- зависимой (NADPH-оксидаза, миелопероксидаза, каталаза), не зависящей от кислорода (лизоцим, катепсины, эластаза, аргиназа, протеазы и другие гидролазы), и в генерации оксида азота (индуцибельная NO-синтаза). Процессы, осуществляемые с участием этих ферментов, описаны далее (см. раздел 2.3.5.2).
Моноциты и макрофаги секретируют некоторые из указанных выше ферментов, а также цитокины, гормоны [адренокортикотропный (АКТГ) и соматотропный гормоны, p-эндорфин и др.], катионные белки, протео- гликаны, метаболиты арахидоновой кислоты, компоненты комплемента, белки межклеточного матрикса (фибронектин, тромбоспондин). Некоторые из них (три последние группы факторов, некоторые ферменты) моноци- ты/макрофаги секретируют спонтанно, но активация обычно усиливает их выработку. С секреторной активностью связано выполнение макрофагами других функций: поставки ряда гуморальных факторов врожденного иммунитета, иммунорегуляторной роли, а также участия в обмене липидов и формировании межклеточного матрикса. Особенность моноцитов/мак- рофагов — быстрая реакция на действие стимулирующих молекул, реализуемая обычно в пределах 1 ч после контакта с молекулой. Однако в этой функции макрофаги значительно уступают нейтрофилам.
В связи со значительными различиями свойств моноцитов/макрофагов и нейтрофилов (см. табл. 2.3) физиологическая роль этих клеток практически не перекрывается, даже несмотря на то, что основная функция тех и других — фагоцитоз. Если нейтрофилы ответственны за самый ранний этап защиты, осуществляемой с помощью фагоцитоза (проходит интенсивно, но кратковременно), то моноциты/макрофаги, помимо фагоцитоза (реализуется менее интенсивно и более продолжительно), выполняют многочисленные другие функции, в том числе опосредованные секретируемыми ими гуморальными продуктами.
Среди секреторных продуктов макрофагов наиболее важную роль в развитии воспаления и реакций врожденного иммунитета играют цитокины. Их секреция, как правило, происходит при активации клеток. Спектр цитокинов, секретируемых моноцитами и макрофагами, очень широк: цитокины семейства IL-1 (IL-1p, IL-18, в меньшей степени IL-1a, представленный на мембране макрофагов и рецепторный антагонист IL-1) и другие провоспалительные цитокины — TNFa, IL-6, IL-12, IL-23, IL-27. Макрофаги продуцируют все 3 разновидности колониестимулирующих факторов (GM-CSF, G-CSF и M-CSF), интерфероны (особенно IFNa, но также IFNp и IFNy), гомеостатический цитокин IL-15, супрессорные цитокины (IL-10 и трансформирующий фактор роста в — TGFP), ростовые/ ангиогенные факторы (фибробластный — FGF, тромбоцитарный — PDGF и сосудистый эндотелиальный — VEGF). Моноциты/макрофаги образуют большую часть провоспалительных хемокинов: CXCL8 (IL-8), CCL5 (RANTES), макрофагальные воспалительные белки (С^3, С^4, ССL9, ССL10, ССL15, ССL18, С^23), макрофагальные хемотаксические белки (ССL2, ССТ7, ССL8, ССL12, ССL13) и др.
Секреторная функция моноцитов и макрофагов (в отличие от грануло- цитов) реализуется в основном по классическому механизму, зависимому от аппарата Гольджи, тогда как дегрануляция, т.е. выброс содержимого лизосом и фаголизосом, играет незначительную роль. Эти формы секреторного процесса отличаются в зависимости от наличия интактных микротрубочек — разрушение микротрубочек колхицином нарушает процесс дегрануляции, но может даже усилить аппарат Гольджи-зависимую секрецию. Дегрануляцией осуществляется выброс продуктов окислительного взрыва, производных оксида азота, кислых гидролаз и других лизосомальных ферментов. Эти факторы обусловливают внеклеточный цитолиз и переваривание клеток и их компонентов, т.е. эффекторные функции врожденного иммунитета, тогда как продукты классического секреторного процесса в большей степени участвуют в регуляции воспаления и реакций врожденного иммунитета. Продукты секреции макрофагов, важные для функционирования врожденного иммунитета, рассмотрены в разделе 2.3.6.
Есть свидетельства неоднородности популяции моноцитов. На основе мембранного фенотипа и функциональных особенностей выделяют две основные разновидности этих клеток: CD14hi CD16- и CD14+ CD16+. Клетки первого типа составляют большинство моноцитов крови. Они имеют более крупные размеры и более высокую плотность, чем вторые клетки. Клеткам с фенотипом CD14hi CD16- свойственна высокая фагоцитарная и бактерицидная активность. Они секретируют полный спектр провоспалительных цитокинов. CD14hi CD16- клетки экспрессируют в большом количестве FcyRI (CD64), рецепторы для хемотаксических факторов и в2-интегрины, особенно Мас-1 ^DUb/CD^). Таким образом, эти клетки имеют необходимые маркеры для эмиграции в очаги воспаления, осуществления фагоцитарной и цитолитической активности и поэтому рассматриваются как предшественники воспалительных макрофагов. Клетки фенотипа CD14+ CD16+ экспрессируют большое количество МНС-II и костимулирующих молекул, обладают относительно слабой фагоцитарной активностью, но эффективно презентируют антиген Т-лимфоцитам, и секретируют IFNa. CD14+ CD16+ клетки рассматривают в качестве предшественников резидентных макрофагов.
Миграция моноцитов в ткани сопровождается их превращением в разнообразные формы макрофагов и дендритных клеток (рис. 2.7, табл. 2.5). Дифференцировка моноцитов в дендритные клетки будет рассмотрена в следующей главе. Выделяют две основные разновидности макрофагов — резидентные и воспалительные (табл. 2.6). Резидентные макрофаги возни-
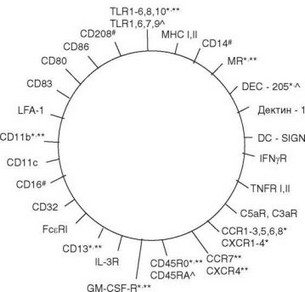
Рис. 2.7. Основные мембранные молекулы дендритных клеток. * Незрелые миелоидные дендритные клетки. ** Зрелые миелоидные клетки. л Плазмоцитоидные дендритные клетки. # Клетки Лангерганса
Таблица 2.5. Маркерные рецепторы и продукты дендритных клеток и моноцитов
|
Молекулы |
Моноциты |
Незрелые миелоидные дендритные клетки |
Зрелые миелоидные дендритные клетки |
Плазмо- цитоидные дендритные клетки |
Зрелые лимфоидные дендритные клетки |
|
TLR |
1, 2, 3, 4, 5, 6, 7, 8, 9, 10 |
1, 2, 3, 4, 5, 6, 8, 10 |
1, 2, 3, 4, 5, 6, 8, 10 |
1, 6, 7, 9, 10 |
1, 2, 4, 6, 7, 9, 10 |
|
Рецепторы цитокинов |
IL-3R+ GM-CSFR++ |
IL-3R+ GM-CSFR++ |
GM-CSFR+ |
IL-3R+++ GM-CSFR+ |
IL-3R++ |
|
CD45R |
RA |
R0 |
R0 |
RA |
RA |
|
CD13/14 |
CD13+ CD14+ |
CD13+ CD14- |
CD13+ CD14- |
CD13- CD14- |
CD13- CD14- |
|
CD11b/c |
CD11b+ CD11c+ |
CD11b+ CD11c+ |
CD11b+ CD11c+ |
CD11b- CD11c- |
CD11b- CD11c+ |
|
Цитокины |
IL-12++ |
IL-12++ |
IL-12-, IFNa/p- |
IFNa/p+++ |
IL-12++, IFNa/p+ |
|
Хемокино- вые рецепторы |
|
CCR1, CCR2, CCR5, CCR6, CXCR1, CXCR2, CXCR3, CXCR4 |
CCR7, CXCR4 |
|
|
Таблица 2.6. Сравнительная характеристика резидентных и воспалительных макрофагов
|
Показатель |
Резидентные макрофаги |
Воспалительные макрофаги |
|
Направление миграции |
В нормальные ткани |
В воспалительные ткани |
|
Хемокиновые рецепторы |
CCR5, CXCR4, CX3CR1 |
CCR1, CCR2, CCR4, CCR7, CXCR1, CXCR2, CXCR4, CX3CR1 |
|
Fcy-рецепторы |
СD16+ CD32+ CD64± |
CD16- CD32++ CD64+ |
|
Экспрессия CD14 |
CD14+ |
CD14+++ |
|
Экспрессия CD62L |
CD62L- |
CD62L++ |
|
Cрок жизни |
Годы |
Недели |
|
Функция |
Тканевой гомеостаз, клиренс апоптотических клеток |
Развитие воспаления, защита от патогенов, заживление ран |
Примечание. - — нет экспрессии; + — слабая экспрессия; ++ — сильная экспрессия;
— очень сильная экспрессия
кают в результате спонтанной («плановой») миграции моноцитов из кровотока в ткани, не связанной с воспалением, тогда как воспалительные макрофаги образуются в процессе экстренной миграции в очаги воспаления. Превращение в макрофаги сопровождается увеличением размера и формы клеток (обусловлены перестройкой цитоскелета), изменением экспрессии некоторых мембранных молекул (ослабевает экспрессия CD13, CD14, CD15, Pj-интегринов, FcyRI, усиливается экспрессия CD16). Это сказывается на ответе клеток на внешние стимулы. Воспалительные макрофаги обладают высокой фагоцитарной и бактерицидной активностью, выделяют ряд цитокинов и других гуморальных веществ, важных для формирования воспаления и реализации иммунной защиты. Эти свойства позволяют воспалительным макрофагам играть роль эффекторных клеток воспаления и врожденного иммунитета. Резидентные макрофаги выполняют преимущественно гомеостатические и регуляторные функции, участвуя в разрушении старых клеток и регуляции иммунных и воспалительных процессов, а также выступают в роли АПК. Резидентные макрофаги обладают более длительным сроком жизни (годы по сравнению с неделями для воспалительных макрофагов).
Воспалительные и резидентные моноциты мигрируют из кровотока в ткани по-разному, поскольку в первом случае решающая роль принадлежит гуморальным факторам и молекулам адгезии, индуцируемым в процессе воспаления, а во втором — невоспалительным гомеостатическим факторам. В качестве хемокина для резидентных макрофагов выступает фракталкин (GX3CL1). Рецепторы для этого хемокина (CX3CR1) экспрессированы в большом количестве преимущественно на CD14+CD16+ моноцитах, при миграции в ткани становящихся резидентными макрофагами. OD14hiCD16- моноциты слабо экспрессируют этот рецептор, но несут рецептор CCR2 для провоспалительных хемокинов преимущественно группы МСР (CCL2, CCL7, CCL8, CCL13, CCL16). В связи с этим СD14hiCD16- клетки обладают способностью мигрировать в очаги воспаления и превращаться в воспалительные макрофаги. Наличие рецептора CCR2 — очень важное свойство СD14hiCD16- клеток, поэтому эту субпопуляцию моноцитов иногда обозначают как CD14hlCCR2+. Эти клетки экспрессируют еще несколько хемокиновых рецепторов, отсутствующих у предшественников резидентных макрофагов: CCR1, CCR4, ССR7, CXCR1, CXCR2. Эти рецепторы распознают практически все провоспалительные и часть гомеостатических цитокинов. В свою очередь, предшественники резидентных макрофагов, помимо рецептора для фракталкина, несут еще несколько хемокиновых рецепторов, отсутствующих или слабо экспрессированных на воспалительных моноцитах — ССR5 и CXCR4 (отметим, что эти рецепторы служат корецепторами для ВИЧ и, следовательно, способствуют инфицированию этим вирусом макрофагов).
Резидентные макрофаги, локализованные в разных органах, могут существенно различаться по морфологии, составу экспрессируемых поверхностных маркеров, спектру секретируемых цитокинов и функциям. Большинство из них имеют собственные названия. Так, макрофаги печени, называемые клетками Купфера, имеют звездчатую форму; они занимают пространство между сосудами печени и гепатоцитами и участвуют в фильтрации продуктов, поступающих из кровотока в паренхиму печени. Численность этих клеток очень велика: на их долю приходится до 50% клеток мононуклеарной фагоцитирующей системы. Определенным своеобразием отличаются альвеолярные макрофаги (способны мигрировать в просвет альвеол), перитонеальные макрофаги, макрофаги центральной нервной системы (микроглия), почек (мезангиальные клетки), костей (остеокласты), тимуса (их важнейшая функция состоит в удалении тимоцитов, в массовом порядке погибающих в процессе развития и селекции), макрофаги вторичных лимфоидных органов и т.д. Вариабельность макрофагов проявляется также и на уровне активированных клеток. Однако в этом случае разнообразие обусловлено не только собственными свойствами моноцитов/макрофагов, но и природой стимуляторов.