Дофаминергическая система
Дофамин, как и норадреналин и адреналин, относится к группе катехоламинов. Будучи независимым нейромедиатором в ЦНС, он еще и предшественник норадреналина во всех центральных и периферических нейронах. Синтез дофамина происходит в аксоплазме. Образовавшийся дофамин депонируется в везикулах. После выброса в синаптическую щель (экзоцитоз) дофамин, как и норадреналин, разрушается с помощью ферментов: моноами- ноксидазы (МАО) и катехол-О-метилтрансферазы (КОМТ). В дофаминергических нейронах и их аксонах преобладает МАОв. Часть неразрушившегося медиатора захватывается специальной транспортной системой, переносится в терминаль и может быть использована повторно.
Дофаминергические нейроны и пути. В целом дофаминергические нейроны сконцентрированы в заднем мозге (группа ядер А8-А10), большинство их обнаружено в компактной зоне черного вещества мозга человека — черной субстанции (Ад).
В дофаминергической системе мозга различают 7 отдельных подсистем (систем, трактов), основные — нигростриатная, мезокорти- кальная, мезолимбическая и тубероинфундибулярная. Тела нейро- 60
переднего мозга
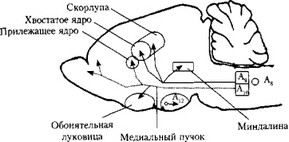
Рис. 11. Дофаминергические ядра и основные дофаминергические пути (цит. по: Webster R. А., 2001):
д8_д9 _ черная субстанция; А10 — вентральное поле покрышки среднего мозга; А12 — аркуатное ядро гипоталамуса
нов нигростриатной, мезокортикальной и мезолимбической систем образуют комплекс нейронов черной субстанции и вентрального поля покрышки (рис. 11). Они составляют непрерывную клеточную сеть, проекции которой частично перекрываются, поскольку аксоны этих нейронов идут вначале в составе одного крупного тракта (медиального пучка переднего мозга), а оттуда расходятся в разные мозговые структуры. Формирование нигростриатной, мезолимбической и мезокортикальной систем определяется областями, где оканчиваются аксоны дофаминергических нейронов, т.е. локализацией их проекций.
Нигростриатная система. Нигростриатный тракт — самый мощный в дофаминергической системе мозга. Аксонами нейронов этого тракта выделяется около 80% мозгового дофамина. Тела дофаминергических нейронов, образующих этот путь, находятся в основном в компактной части черной субстанции (А9), хотя незначительное число тел нейронов было обнаружено и более вентрально (А8). Аксоны нейронов Ag—Ад, а также часть волокон от нейронов латерального отдела вентрального поля покрышки среднего мозга образуют нигростриатный тракт. Тела нейронов черной субстанции у человека и приматов содержат гранулы липопротеинового пигмента — нейромеланина, придающего черный цвет нейронам этого ядра.
Наиболее плотно расположены дофаминергические волокна в неостриатуме. Они начинаются от латеральных отделов черной субстанции того же полушария и оканчиваются на нейронах хвостатого ядра и скорлупы. Дофаминергическую иннервацию получают так- же другие структуры, в частности, бледный шар и субталамическое ядро. В хвостатом ядре более плотная иннервация в головке и значительно меньшая плотность дофаминергических проекций в каудальной части.
Мезокортикалъная система. Тела нейронов, образующих мезокор- ти-кальный тракт, находятся в вентральной части покрышки среднего мозга (А)0), а основные проекции этих нейронов достигают лобной (преимущественно префронтальной, поле 10, по Бродману) коры. Соответствующие окончания расположены в основном в глубоких слоях лобной коры (V—VI). Мезокортикальная дофаминовая система оказывает большое влияние на активность нейронов, образующих корково-корковые, корково-таламические и корково-стриатные пути.
Мезолимбическая система. Тела нейронов этой системы расположены в вентральном поле покрышки среднего мозга и частично в компактной части черной субстанции. Их отростки идут в аккум- бентное (прилежащее) ядро, поясную извилину, энторинальную кору, миндалину, обонятельный бугорок, гиппокамп, парагиппокампальную извилину, перегородку и другие структуры лимбической системы мозга. Имея обширные связи, мезолимбическая система опосредованно проецируется также на лобную кору и гипоталамус. Это определяет широкие функции мезолимбической системы, которая участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
Тубероинфундибулярный тракт образован аксонами нейронов, расположенных в аркуатном ядре гипоталамуса (А12). Отростки этих нейронов достигают наружного слоя срединного возвышения.
Рецепторы. Основные типы дофаминовых рецепторов представлены Д j - и Д2-рецепторами. Выделяют и Д5-рецепторы как разновидность Д,-рецепторов, а также Д3- и Д4-рецепторы как разновидность Д2-формы. Дофаминовые рецепторы относятся к метаботропным и сопряжены с аденилатциклазой, которая участвует в синтезе вторичного мессенджера — цАМФ. Д4- и Д5-рецепторы активируют аденилатциклазу, тогда как Д2-, Д3- и Д4— ее тормозят. Дофаминовые рецепторы могут быть расположены как пост-, так и пресинаптически, контролируя высвобождение медиаторов не только дофаминергической, но норадренер- гической, холинергической медиаторных систем.
Дррецепторы наиболее представлены в мозге человека в стриа- туме (хвостатом ядре и скорлупе), прилежащем ядре и обонятельной луковице, незначительно — в гипоталамусе и коре. Наибольшая плотность Д5-рецепторов обнаруживается в гиппокампе и гипоталамусе.
Д2-рецепторы преобладают в стриатуме, прилежащем ядре и обонятельной луковице. Обнаружены они также на телах нейронов черной субстанции и вентральной тегментальной области, где являются ауторецепторами, регулирующими высвобождение дофамина. В стриатуме Д2-рецепторы расположены на афферентах нигростри- атного тракта, выступая как пресинаптические ауторецепторы, а также на холинергических нейронах. Они увеличивают К+-проводи- мость, вызывая гиперполяризацию холинергических нейронов, а также тормозят вход Са2+ через потенциал-зависимые Са2+-каналы. Поэтому введение агонистов дофамина сопряжено с угнетением выделения ацетилхолина холинергическими нейронами стриатума. В тех случаях, когда Д2-рецепторы в стриатуме функционируют как ауторецепторы, их эффект проявляется в снижении высвобождения дофамина.
Д3-рецепторы менее представлены, чем Д2-, в основном в лимбической системе (прилежащее ядро и обонятельная луковица), а также в гипоталамусе. Незначительное количество обнаружено в хвостатом ядре, а также в качестве ауторецепторов на дофаминергичес- ких нейронах черной субстанции.
Д4-рецепторы очень немногочисленны. Локализованы во фронтальной коре, в среднем мозге и миндалине.
Функции дофаминергической системы в ЦНС. От состояния дофамин- ергической системы зависит центральная регуляция двигательной активности, поведенческие и психические функции, продукция ряда гипофизарных гормонов (пролактин) и функция центра рвоты.
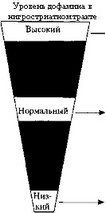
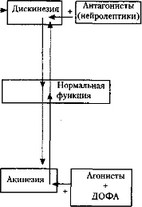
Двигательная активность. У больных паркинсонизмом выявлен недостаток дофамина в ЦНС. Устранить этот недостаток можно с помощью предшественника дофамина — диоксифенилаланина (ДОФА), применение которого уменьшает скованность и акинезию у таких больных (рис. 12). Повышенная активность дофаминергической системы нигростриатного тракта, как при хорее Гентинггона, вызывает повышенную двигательную активность, дискинезию (см. рис. 12). Ее проявления можно контролировать введением антагонистов дофаминовых рецепторов (нейролептиков). Применение дофаминовых антагонистов (хлорпромазин) при лечении шизофрении снижает активность дофаминергической системы, что может вызвать экстрапирамидные нарушения, характерные для болезни Паркинсона.
При внутрижелудочковом введении животным дофамина возникает выраженная моторная гиперактивность, а также характерное стереотипное поведение. Стереотипия вызвана активацией нигро- стриатных путей, так как она исчезает после поражения черной субстанции и появляется вновь после введения в стриатум апоморфи- на, агониста дофаминовых рецепторов.
Психические функции. Существует гипотеза, что причиной психозов является усиление функции дофаминергических синапсов в результате увеличения выброса медиаторов из пресинаптических окончаний, либо повышения чувствительности постсинаптических рецепторов к дофамину, либо активации его синтеза. Определяющую роль дофаминовой гиперактивности в генезе психозов подтверждает клиническое сходство фенаминовой интоксикации с картиной шизофренического психоза, который проявляется различными психотическими симптомами, в частности галлюцинациями. Известно, что центральное действие фенамина (амфетамина) связано в первую очередь с воспроизведением эффектов дофамина. Это указывает на то, что дофамин играет важную роль в контроле и проявлении психотических симптомов. Полагают, что психозы вызываются медиацией дофаминергических мезокортикальных и мезолимбических путей.
награждение и подкрепление. Мы ожидаем награду как удовольствие. У животных это выражается готовностью к поиску или подходом, например, к кормушке или рычагу, нажатие на который вызыует у НИх наслаждение (удовольствие). Если животному после вживления микроэлектродов в структуры, связанные с реализацией Положительного подкрепления, дать возможность произвольно раздражать их электрическим током, то оно будет делать это безостановочно в течение длительного времени, вплоть до полного истоц^ния. Считают, что участки мозга и нейромедиаторы, вызывающие самостимуляцию, могут «отвечать» и за естественные проявление удовольствия и вознаграждения. К таким участкам относят область медиального пучка переднего мозга, включающего аксоны норадренергических нейронов голубого пятна и дофаминергических нейр^ов, расположенных в вентральной тегментальной области. До- фамидергические нейроны группы ядер А10 иннервируют аккумбен- тное Пдро и префронтальную кору. При самостимуляции через электроду имплантированные в вентральную тегментальную область, выявден0 повышенное высвобождение дофамина в этих структурах. Порор электрического раздражения, вызывающего самостимуляцию, снижался при введении амфетамина и повышался при введении антагонистов дофаминовых рецепторов.
Т\ким образом, выделение дофамина в аккумбентном ядре ассоциируется с процессами подкрепления и вознаграждения. Однако резу^таты этих экспериментов не позволяют сделать вывод, можно ли связывать усиление дофаминергической медиации в этом ядре с осознанием вознаграждения или с запуском моторного акта, необходимого для получения вознаграждения.
Источник: Белова Е. И., «Основы нейрофармакологии» 2006