Норадренергическая система
Норадренергические нейроны и пути. Тела норадренергических нейронов в ЦНС млекопитающих находятся (рис. 10) в стволе мозга (А,—А?), главным образом в мосте (голубое пятно), в продолговатом мозге (латеральные тегментальные ядра и ядро одиночного тракта). Наиболее крупными являются голубое пятно (Ag — locus coeruleus, А( — subcoeruleus) и латеральные тегментальные ядра (Aj, А3, А5, А7). Многочисленные области мозга иннервируются волокнами нейронов голубого пятна и латеральной тегментальной области, при этом фронтальная кора, гиппокамп и обонятельные луковицы получают проекции исключительно от нейронов голубого пятна, в то время как большинство

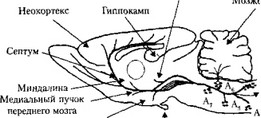
гипоталамических ядер иннервируются нейронами латеральной
тегментальной области.
Голубое пятно представляет наибольший интерес, так как оно содержит около 45% норадренергических нейронов мозга. Многочисленные нейроны голубого пятна образуют диффузные проекции большой протяженности, достигающие практически всех отделов ЦНС: коры больших полушарий, лимбической системы, таламуса, гипоталамуса, спинного мозга. На активность этих нейронов влияют ГАМК-ергическая тормозная система и возбудительная глутамат - ергическая система. Большинство волокон от нейронов голубого пятна проецируются ипсилатерально. Плотность иннервации варьирует в различных регионах мозга. При этом 50% нейронов голубого пятна иннервируют кору головного мозга и мозжечок.
Рецепторы. В ЦНС имеются как а-, так и p-адренорецепторы. Обнаружены а,-адренорецепторы во многих отделах мозга, и их распределение неодинаково в разных регионах. В голубом пятне доминируют а2-адренорецепторы, и этот подтип рецепторов ответственен за сокращение высвобождения медиатора в синаптическую щель, а также за снижение нейрональной активности целевых клеток, что является результатом уменьшения образования цАМФ, увеличения К+-токов, уменьшения Са2+-токов.
В противоположность а2-адренорецепторам p-адренорецепторы активируют синтез цАМФ. В мозге Р,- и Р2-рецепторы имеют раз-
личное распределение. Почти 60% p-адренорецепторов неокортекса относится к Pj-подтипу, в то время как в мозжечке доминирует преимущественно Р2-подтип. Активация p-адренорецепторов пирамидных клеток коры приводит к увеличению их возбудимости, а в таламических релейных нейронах вызывает деполяризацию и способствует улучшению проводимости входных афферентных сигналов через релейные ядра таламуса. Однако в целом трудно определить электрофизиологические изменения, возникающие в результате активации p-адренорецепторов. Это связано с неопределенностью их локализации. Так, p-адренорецепторы в неокортексе могут располагаться на интернейронах, и их активация сопровождается тормозными процессами; или обеспечивать прямое аксосоматическое взаимодействие с целевыми клетками, в результате развиваются процессы возбуждения. Кроме того, эффекты возбуждения этих рецепторов зависят от уровня тонической активации целевых клеток. В целом полагают, что возбуждение р-адренорецепторов в ЦНС приводит к увеличению возбудимости и реактивности целевых клеток, что вносит существенный вклад в поддержание уровня активации мозга и обеспечение селективного внимания.
Функции норадренергической системы в ЦНС. Считается, что в ЦНС норадреналин участвует в процессах регуляции функционального состояния, селективного внимания, оказывает влияние на двигательную активность, мотивационное и эмоциональное поведение, обеспечивает вегетативные проявления большинства эмоциональных реакций. Влиянием на синтез, хранение и обратный захват норадреналина объясняется центральное действие многих нейротропных веществ. Так, повышение активности норадренергической системы, обусловленное введением L-ДОФА, предшественника норадреналина, сопровождается повышением уровня двигательной активности, улучшением выработки условной реакции активного избегания болевого наказания ударом электротока. Понижение активности норадренергической системы мозга вследствие блокады синтеза норадреналина с помощью дисульфирама резко снижает все показатели поведенческого возбуждения, кроме того, существенно замедляется или становится невозможной выработка условной реакции активного избегания.
Регуляция уровня бодрствования. Так как норадренергические пути в ЦНС имеют диффузное распределение, а синаптические эффекты норадреналина довольно разнообразны, полагают, что активность норадренергической системы оказывает влияние на уровень
активации (arousal) ЦНС. Стимуляция голубого пятна повышает игательную активность животного и вызывает ЭЭГ-десинхрони аяпию в то время как поражение норадренергических нейронов неи- потоксином ведет к ЭЭГ-синхронизации, увеличению длительности медленноволнового сна и уменьшению парадоксального сн /таFM-сна) Снижение активности нейронов голубого пятна предшествует началу сна. Некоторые (REM-off) нейроны прекращают свою активность при REM-сне. Полагают, что изменение активности норадренергических нейронов голубого пятна играет «разреша- ющую» роль в начало сна.
С адренергической системой связаны преимущественно стиму- тирующие влияния на многие функции мозга. Это имеет отношение не только к циклу сонбодрствование, но и к более специфическим процессам, в частности, к селективному вниманию и бдительности.
Регуляция селективного внимания. В электрофизиологических исследованиях активность одиночных нейронов голубого пятна увеличивалась под действием сенсорных стимулов. Эта реакция была наиболее выражена на стимулы, вызывающие физическии дискомфорт, на интероцептивные стимулы и некоторые возбуждающие внешние сигналы. Поэтому предположили, что центральные норадрен- ергические нейроны — часть «системы настораживания», их нейро нальная активность снижается на повторяющееся предъявление тестового стимула, что лежит в основе поведенческого привыкания. Новизна - тот ключевой фактор, который увеличивает норадренер- гическую передачу, вызывая изменение селективного внимания. 1ак, при предъявлении нейтрального стимула (звонка) во фронтальной коре мозга крысы наблюдалось высвобождение норадреналина толь ко в том случае, если ее помещали в новые условия, в яРу освещен ное место. Вероятно, норадренергические нейроны голубого пятна, иннервирующие эту область, отвечают усилением активности в ответ на изменение внешних стимулов. ^
Усиление активности норадренергических нейронов, обеспечивающих процессы селективного внимания, носит характер фазичес- ких ответов. Эти ответы зависят от тонической активации нейронов голубого пятна, которая определяется уровнем arousal. При этом оптимальные фазические ответы наблюдаются только на среднем уровне тонической активации нейронов.
Регуляция эмоционального состояния. В изменении норадренер- гической передачи существенную роль играет эмоциональная составляющая предъявляемого стимула, а норадренергическая система мозга принимает непосредственное участие в формировании эмоци-
онального состояния. Недостаточность норадренергической передачи приводит к депрессии, в то время как ее умеренная активность необходима для формирования интереса при выполнении когнитивных задач. Чрезмерная норадренергическая медиация отмечается в состоянии тревоги и смятения. Между плотностью постсинаптических p-адренорецепторов в коре мозга крысы и устойчивостью животных к действию мягкого стрессирующего стимула существует положительная корреляция. При усилении стресса корреляционная связь между ними становится отрицательной. Норадренергическая система мозга принимает участие и в формировании эмоции ярости и агрессии. Повышение агрессивности животных связывают с ростом концентрации норадреналина в структурах мозга и ослаблением тормозного влияния серотонина.
Таким образом, для формирования адекватного ответа на внешнее воздействие необходимо специфическое изменение норадренергической передачи. В формировании поведенческой реакции на стимулы внешней среды участвует не только норадренергическая, но и другие нейромедиаторные системы.
Источник: Белова Е. И., «Основы нейрофармакологии» 2006