
Аутоиммунные заболевания продолжают оставаться одной из самых сложных проблем клинической иммунологии.
Вот уже почти сто лет одной из главных догм иммунологии, сформулированной Паулем Эрлихом, является представление о том, что в норме иммунная система не должна развивать иммунный ответ против собственных тканей, поскольку это может привести к гибели организма. П. Эрлих назвал это "ужасом самоотравления" ("horror autotoxi- cus"). В настоящее время этот феномен известен под названием "иммунная толерантность", которая развивается в эмбриональном и раннем постнатальном периоде и заключается в том, что в организме создаются условия, при которых иммунная система не реагирует с аутоантигенами (self-антигенами) (об этом уже упоминалось в соответствующей главе).
Таким образом, аутоиммунитет характеризуется потерей (нарушением, исчезновением) толерантности, или естественной неотвечаемо- сти по отношению к собственным антигенам. Как следствие, продуцирующиеся аутоантитела и/или цитотоксические клетки приводят к развитию заболевания.
Однако способность иммунной системы распознавать аутоантиген не всегда несет патологический потенциал. Так, например, распознавание собственных молекул главного комплекса гистосовместимости при реализации иммунного ответа, антиидиотипический ответ против self-идиотипов и др.; все это позволяет иммунной системе выполнить свою основную функцию иммунного надзора.
В настоящее время описано очень большое количество аутоиммунных заболеваний. Предполагается, что иммунная Система при соответствующих условиях может развить иммунный ответ против любого аутоантигена.
Аутоиммунные заболевания делят на две группы:
- органоспецифические — например тяжелая миастения, тиреоидит Хашимото, болезнь Грейвса (тиреотоксикоз с диффузным зобом) и др.;
- системные (неорганоспецифические) — например системная красная волчанка, ревматоидный артрит и др.
Аутоиммунные заболевания (неполный перечень, призванный показать встречаемость этой патологии практически во всех медицинских специальностях)
- Системная красная волчанка
- Ревматоидный артрит
- Склеродермия
- Дерматополимиозит
- Смешанные болезни соединительной ткани
- Синдром Шегрена (сухой синдром)
- Псориаз
- Витилиго
- Дерматит герпетиформный
- Пузырчатка обычная
- Буллезный пемфигоид
- Болезнь (синдром Рейтера)
- Болезнь Бехтерева
- Рассеянный множественный склероз
- Острый (пост-)инфекционный полиневрит (синдром Гийена — Барре)
- Тяжелая миастения
- Тиреоидит Хашимото (аутоиммунный) .
- Болезнь Грейвса (тиреотоксикоз с диффузным зобом)
- Сахарный диабет инсулинзависимый (I типа)
- Аутоиммунное поражение надпочечников (болезнь Аддисона)
- Аутоиммунная полиэндокринопатия
- Саркоидоз
- Идиопатический легочный фиброз
- Неспецифический язвенный колит
- Болезнь Крона (региональный энтерит)
- Аутоиммунный гастрит, тип А
- Первичный биллиарный цирроз
- Хронический активный гепатит
- Аутоиммунная энтеропатия
- Целиакия (глютенчувствительная энтеропатия)
- Гломерулонефрит
- Синдром Гудпасчера
- Аутоиммунный орхит
- Аутоиммунное бесплодие
- Первичный синдром антифосфолипидных антител -
- Аутоиммунный увеит
- Симпатическая офтальмия
- Аутоиммунный конъюнктивит
- Узловатый полиартериит
- Гигантоклеточный гранулематозный артериит (ревматическая полимиалгия)
- Пернициозная анемия
- Аутоиммунная гемолитическая анемия
- Аутоиммунная тромбоцитопения
- Аутоиммунная нейтропения и др.
В одних случаях развитие аутоиммунитета (срыв толерантности) может быть первичным и служить причиной развития заболевания, в других, особенно при длительных хронических заболеваниях (например хронический пиелонефрит, хронический простатит и др.), — вторичным. и являться следствием заболевания, замыкая "порочный круг" патогенеза.
Нередко у одного и того же больного развивается несколько аутоиммунных заболеваний, особенно это относится к аутоиммунным эн- докринопатиям.
Аутоиммунные заболевания часто ассоциируются с лимфоидной гиперплазией, злокачественной пролиферацией лимфоидных и плазматических клеток, иммунодефицитными состояниями — гипогамма- глобулинемией, селективным дефицитом IgA, недостаточностью компонентов комплемента и др. Системные аутоиммунные заболевания часто развиваются в зрелом возрасте.
В настоящее время предложено около двух десятков теорий, объясняющих причины срыва толерантности и, как следствие, развития аутоиммунитета. Приведем основные из них.
- Теория "запретных" клонов. Известно, что при индукции толерантности на определенных этапах развития (созревания) иммунной системы происходит элиминация (разрушение) тех Т- и В-лимфоци- тов, которые обладают аутореактивностью — способностью реагировать с ауто (self) — антигенами. Согласно теории "запретных" клонов, по тем или иным причинам в тимусе и костном мозге не происходит полная элиминация аутореактивных Т- и В-лимфоцитов, что в будущем, при стечении определенных обстоятельств, может привести к срыву толерантности.
- Теория секвестрированных (забарьерных) антигенов. Известно, что определенные ткани ограждены гистогематическими барьерами (половые железы, ткани глаза, мозга, щитовидной железы и др.). В связи с этим при созревании иммунной системы антигены таких тканей не контактируют с лимфоцитами и не происходит элиминации соответствующих клонов клеток. При нарушении гистогематического барьера и попадании антигенов в кровоток собственные иммунокомпетентные клетки распознают их как чужеродные и запускают весь механизм иммунного ответа.
- Теория расстройства иммунологической регуляции.
- Снижение функции Т-лимфоцитов-супрессоров. Считается, что Т-лимфоциты-супрессоры подавляют способность В-лимфоцитов продуцировать антитела против собственных тканей, поддерживая таким образом состояние толерантности. При снижении количества или функции Т-супрессоров потенциально аутореактивные В-клетки начинают реагировать на собственные тканевые антигены, а появляющиеся аутоантитела приводят к развитию аутоиммунного заболевания.
- Нарушение функции Т-лимфоцитов-хелперов. В частности, при ее повышении могут создаваться условия, благоприятные для инициации ответа со стороны аутореактивных В-лимфоцитов на ¦ собственные антигены, даже при нормальной функции Т-супрессоров. Таким образом, потенциальные возможности развития аутоиммунитета, имеющиеся в организме, реализуются за счет нормально функционирующих иммунологических регуляторных механизмов, включающих, прежде всего, Т-лимфоциты — супрессоры и хелперы.
- В последние годы все большую популярность приобретает гипотеза, согласно которой в основе аутоиммунной патологии лежат расстройства иммунной регуляции, обусловленные нарушением продукции соответствующих цитокинов Т-лимфоцитами-хелперами I и Шипов.
- Теория нарушения идиотип-антиидиотипических взаимодействий.
Современные модели иммуного ответа предполагают, что иммунная система обладает саморегулировкой и может реагировать на свои собственные продукты с последующей супрессией или стимуляцией этой реакции. Известно, что в сыворотке крови больных и здоровых лиц можно обнаружить антитела против собственных Ig (первым антителом такого типа, обнаруженным у человека, был ревматоидный фактор). Идиотипическая детерминанта (идиотип) тесно связана с индивидуальной структурой активного центра молекулы Ig. Вначале считалось, что продукция аутоантител против собственных Ig — результат нарушения процесса распознавания "своего", и это является либо причиной, либо симптомом заболевания. Однако впоследствии многие исследователи обнаружили антииммуноглобулины в сыворотке крови здоровых лиц, исходя из чего предположили, что продукция антииммуноглобулинов представляет собой физиологический, а не патологический процесс. На этой основе была разработана модель иммунной системы, в которой контрольно-регуляторные влияния зависят от множества взаимодействующих компонентов, а антииммуноглобулины, направленные против активного центра молекулы специфического антитела (антиидиотипические антитела) играют ведущую роль. Было сделано предположение (N. К. Erne, 1974), что распознавание идиотипических детерминант и развитие антиидиотипического иммунного ответа представляет собой центральный механизм контроля и регуляции биосинтеза антител. Эта теория получила название сетевой теории регуляции иммунного ответа. .
В теории Ерне можно выделить два основных положения:
- Иммуноглобулины, а также иммуноглобулиновые рецепторы на поверхности антигенреактивных Т- и В-лимфоцитов имеют детерминанты, обладающие (ауто-) антигенными свойствами, и получившие название "идиотип" (идиотипические детерминанты);
- В организме предсуществуют лимфоциты, способные в норме распознать своими рецепторами идиотипические детерминанты и реализовать антиидиотипический ответ. Антиидиотипическое антитело также может быть распознано и на него вырабатываются анти-анти- идиотипические антитела до тех пор, пока иммунный ответ не угаснет. Полагают, что идиотип и анти-антиидиотип являются идентичными структурами.
Исследования последних лет подтверждают важную роль идиотип- антиидиотипических взаимодействий в регуляции иммунного ответа. Необходимо выделить следующие основные положения:
- Антиидиотипический ответ развивается одновременно с обычным иммунным ответом на чужеродные антитела;
- Идиотип-антиидиотипические взаимодействия обусловливают возможность как стимуляции, так и супрессии лимфоцитов под влиянием антиидиотипических антител. С учетом этих данных, становится ясно, что развивающийся одновременно с обычным иммунным ответом антиидиотипический ответ, стимулируя или угнетая первый в зависимости от тех или иных обстоятельств, обеспечивает его саморегуляцию по типу обратной связи.
Таким образом, при осуществлении иммунного ответа развиваются ¦ антитела, иммунные комплексы и/или клеточно-опосредованный иммунный ответ. Для того, чтобы сбалансировать эти медиаторы иммунопатологии и не дать им "работать" против собственных тканей, одновременно включается регуляторный механизм, представляющий собой сложную сеть Т-, В-клеток и антител, координированный как антиидиотипический иммуный ответ. Этот механизм обеспечивает контроль, необходимый для предотвращения повреждения органов- мишеней во время бесчисленных иммунных реакций, генерируемых "хозяином" в пределах собственного организма.
Из сказанного ясно, что нарушение идиотип-антиидиотипических взаимодействий будет способствовать развитию аутоиммунных заболеваний.
- Теория поликлональной активации В-лимфоцитов. Обнаружено, что многие вещества химической или биологической природы обладают способностью индуцировать активацию В-лимфоцитов, которая приводит к их пролиферации и продукции антител. Как правило, такие антитела относятся к иммуноглобулинам класса М. В том случае, если поликлональной активации подверглись аутореактивные В-лим- фоциты, продуцирующие аутоантитела, возможно развитие аутоиммунного заболевания.
Поликлональные активаторы В-лимфоцитов
Липополисахарид
Очищенный белок туберкулина
Протеин A Staphylococcus aureus
Белок, ассоциированный с липидом А .
Т-клеточные и макрофагальные лимфокины Fc-фрагмент Ig
Протеолитические ферменты (например трипсин)
Полианионы (например, декстрана сульфат)
Антибиотики, (например, нистатин, амфотерицин В)
Микоплазма
Некоторые вирусы и вирусные компоненты (вирус Эпштейна — Барра, вирус кори, белки теплового шока)
Паразиты (Trypanosoma cruzi, Plasmodium malariae).
- - Теория развития аутоиммунитета под влиянием суперантигенов.
Бактериальные суперантигены получили свое название в связи со способностью активировать большое количество Т- и В-лимфоцитов независимо от антигенной специфичности этих клеток. Выше упоминалось, что при классическом варианте антигенного распознавания Т-хелпер активируется под влиянием взаимодействия Т-клеточного антигенрас- познающего рецептора (ТАГРР) и пептида, который презентируется антигенпредставляющей клеткой (АПК) в ассоциации с молекулой главного комплекса гистосовместимости класса II (рис. 19). При этом только один (или несколько)- Т-лимфоцитов-хелперов могут быть активированы.
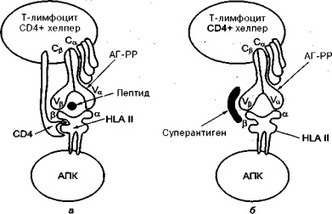
Рис. 19. Активация Т-лимфоцитов-хелперов под влиянием суперантигена: а — классический путь активации; 6 — активация суперантигеном.
Активация Т-лимфоцитов-хелперов под влиянием суперантигенов происходит совсем по-другому. В этом случае суперантиген не поглощается антигенпредставляющей клеткой и не подвергается обычному перевариванию (процессингу) с образованием пептида. При этом суперантиген как бы обходит этот необходимый для специфического распознавания этап и неспецифически связывается с вариабельной частью бета-цепи Т-клеточного распознающего рецептора вне его антигенспецифической зоны (сайта). Происходит своеобразное перекрестное связывание молекул главного комплекса гистосовместимости антигенпрезентирующей клетки с Т-клеточным распознающим рецептором. В случае такого механизма активации Т-лимфоцитов-хелпе- ров возможна одновременная активация большого их количества.
Таким образом, отличительные особенности стимуляции Т-лимфо- цитов под влиянием суперантигенов заключаются в следующем:
- Для этого нет необходимости в переваривании (процессинге) антигена в антигенпредставляющей клетке;
- Такая стимуляция не зависит от антигенной специфичности молекул комплекса HLA и Т-клеточного распознающего рецептора;
- Суперантиген способен стимулировать в 103—104 раз больше лимфоцитов, чем процессированный антиген;
- Аллогенный (чужеродный) суперантиген может стимулировать как хелперы (CD4+), так и киллеры (CD8+) Т-лимфоциты;
- Аутологичный (self) суперантиген может стимулировать только Т-лимфоциты-хелперы (CD4);
- Для полноценной стимуляции Т-лимфоцитов чужеродным суперантигеном необходим дополнительный, костимуляционный, сигнал.
Чужеродные суперантигены описаны для' Staphylococcus aureus (энтеротоксины А, В, С и др., токсин, вызывающий синдром токсического шока, эксфолиативные токсины), Streptococcus pyogenes (эритрогенный токсин, токсины А, В, С, D); для Mycoplasmae arthritidis.
Под влиянием этих суперантигенов могут развиваться следующие заболевания (состояния): пищевая токсикоинфекция, синдром токсического шока, синдром чешуйчатой кожи, ревматическая лихорадка, артрит и др.
Установлено также, что некоторые опухолевые вирусы, находящиеся в геноме клетки в форме провируса, могут кодировать продукцию белка, который вызывает стимуляцию Т-лимфоцитов, выступая в качестве суперантигена.
Рассматриваются три возможных механизма участия суперантигенов в развитии аутоиммунных нарушений.
А. Активация аутореактивных Т-лимфоцитов. Доказано, что супер-
антигены могут непосредственно активировать аутореактивные Т-лим- фоциты, которые затем мигрируют в соответствующие ткани и вызывают аутоиммунные нарушения, продуцируя цитокины и/или реализуя свою киллинговую функцию.
Б. Активация аутореактивных В-лимфоцитов. Осуществляется за счет того, что суперантиген связывает молекулы комплекса HLA класса II, имеющиеся на В-лимфоцитах, с молекулой Т-клеточного антиген- распознающего рецептора. В этом случае активация Т-лимфоцитов происходит без специфического распознавания антигена, а неспецифически под влиянием суперантигена. Тем не менее, такой Т-лимфо- цит продуцирует соответствующие цитокины, которые способствуют тому, что активированный аутореактивный В-лимфоцит начинает продуцировать аутоантитела. Последние образуют иммунные комплексы и, оседая в тканях, вызывают их повреждение. Не исключается, что В-лимфоциты могут активироваться и через собственный анти- генраспознающий иммуноглобулиновый рецептор.
В. Активация антигенпредставляющих клеток. Суперантигены могут активировать антигенпредставляющие клетки, например макрофаги. Это приводит к высвобождению из них цитокинов, супероксидных . анионов и других медиаторов воспаления. Активация макрофагов может также привести к нарушению переваривания (процессинга) антигенов с последующей презентацией аутоантигенов аутореактивным Т-лимфоцитам.
- Теория генетической предрасположенности. Согласно современным данным, существует генетически детерминированная предрасположенность к развитию аутоиммунных заболеваний. Эта предрасположенность контролируется по меньшей мере шестью генами, расположенными на разных хромосомах. Часть из них расположена в главном комплексе гистосовместимости (HLA) человека, роль которого . в реализации иммунного ответа является первостепенной. Установлено, что большинство аутоиммунных заболеваний ассоциируются с наличием в HLA-фенотипе человека следующих антигенов: DR2, DR3, DR4 и DR5. Например, ревматоидный артрит ассоциируется с HLA-DR4, тиреоидит. Хашимото — с HLA-DR5, рассеянный множественный склероз — с HLA-DR2, системная красная волчанка — с HLA-DR3.
Доказано также, что аутоиммунные заболевания развиваются намного чаще у женщин, чем у мужчин. Например, частота встречаемости- системной красной волчанки у женщин в 6—9 раз выше, чем у мужчин. Считается, что в данном случае важную роль играют половые гормоны.
В рамках теории генетической предрасположенности выдвинуто несколько гипотез, объясняющих участие продуктов HLA-комплекса в патогенезе заболеваний вообще и аутоиммунных в частности.
- Согласно рецепторной гипотезе, одной из наиболее ранних, определенные HLA-антигены являются рецепторами для вирусов, облегчающими их фиксацию и проникновение в клетку. Эта гипотеза имеет много аргументов как в свою пользу, так и против. Например при таком заболевании явно вирусной этиологии, как полиомиелит, а также при инфекционном мононуклеозе достоверной корреляции с HLA- антигенами не обнаруживается.
Б. Гипотеза о модификации (изменении) аутологичного, своего, антигена (alteredself). Согласно этой гипотезе, модифицированный аутологичный антиген распознается иммунной системой как чужеродный (nonself), что приводит к срыву толерантности.
- Гипотеза о влиянии гипотетического Ir-гена на предрасположенность к заболеваниям (нарушение селекции антигенных детерминант, наличие "дыр" в репертуаре Т-лимфоцитов, нарушение супрессии, опосредованной Т-лимфоцитами).
Г. Гипотеза о влиянии неклассических генов, картирующихся в пределах системы HLA.Например гены HSP-70, TNF, недостаточность С4А, С2 ассоциируются с системной красной волчанкой и пиогенной инфекцией.
- Теория молекулярной мимикрии. Термин "мимикрия" в свое время был предложен для объяснения подобия, идентичности антигенных детерминант некоторых микроорганизмов антигенным детерминантам хозяина, в связи с чем их распознавание иммунной системой не происходит, что и обусловливает развитие инфекционного заболевания. В настоящее время теория молекулярной мимикрии видоизменилась и представлена двумя вариантами (рис. 20).
А. Согласно первому варианту теории, некоторые микроорганизмы действительно обладают перекрестной реактивностью с антигенными детерминантами хозяина возможно не за счет идентичности, а за счет достаточно выраженного подобия (гомологии). Это обстоятельство имеет свое объяснение. Действительно, главнейшая (и, видимо, первоначальная) роль иммунной системы состоит в том, чтобы защитить организм от инфекций. Для этой цели основные клетки иммунной системы — Т- и В-лимфоциты — снабжены антигенраспозна- ющими рецепторами самой разной специфичности, что позволяет им распознать любой, внедрившийся в организм инфекционный агент.
Распознав чужеродный агент, иммунная система защищается двумя основными механизмами: 1) продукцией гуморальных антител;
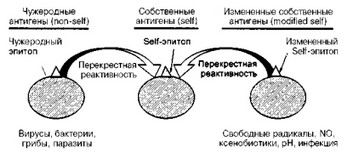
Рис. 20. Роль мимикрии в развитии аутоиммунитета (объяснение в тексте).
- генерацией цитотоксических Т-лимфоцитов. При первом механизме защиты антитела поражают внеклеточные инфекционные агенты и их токсины, образуя иммунные комплексы; при втором механизме — для спасения всего организма цитотоксическим Т-лимфоцитам приходится разрушать собственные клетки, в которых прячутся внутриклеточные возбудители.
Таким образом, иммунитет к инфекционным агентам довольно часто имеет иммунологический компонент либо в виде иммунных комплексов, либо в виде цитотоксических Т-лимфоцитов. Отсюда следует, что, развивая противоинфекционный ответ, иммунная система должна "выбирать" силу, с которой она защищается: ответ должен быть достаточным для элиминации возбудителя, но безвредным для организма. Этот баланс зависит от многих условий: а) степени выраженности и продолжительности инфекции; б) повреждающего действия патогена и степени иммунного ответа; в) количества и значимости тех клеток хозяина, которые были разрушены при попытке элиминации внутриклеточного возбудителя.
Микроорганизмы экспрессируют множество антигенов, которые подобны, если не идентичны, антигенам хозяина. Если бы все Т- и В- лимфоцйты, способные реагировать с этими антигенами, были элиминированы в период создания толерантности, то в защитных способностях иммунной системы были бы большие пробелы, что позволило бы этим микроорганизмам беспрепятственно внедряться в организм. Однако это не так, следовательно, те Т- и В-лимфоциты, которые распознают инфекционные агенты, обладающие антигенами, подобными антигенам хозяина (перекрестно реагирующие антигены), могут реагировать с собственными клетками, т. е. обладают аутореактивностью.
Таким образом, при создании толерантности в эмбриональном и раннем постнатальном периоде полного разрушения аутореактивных Т- и В-лимфоцитов не наступает. Сохранив аутореактивные Т- и В- лимфоциты, организм увеличивает возможности иммунной системы противостоять инфекционным агентам, обладающим подобными антигенными структурами. И как следствие, развитие протективного антиинфекционного иммунного ответа при определенных условиях может приводить к развитию аутоиммунного ответа (табл. 17).
Таблица 17. Примеры молекулярной мимикрии при аутоиммунной патологии у человека
|
Заболевание |
Self антиген |
Non-self антиген |
|
Рассеянный склероз |
Основный белок миелина |
Вирус гепатита В, полимера- раза, фосфолипидный белок Saccharomyces cerevisial, белок CRMI |
|
Инсулинзависимый сахарный диабет |
Декарбоксилаза глутаминовой кислоты |
Coxsackie virus, белок 32-С |
|
Первичный биллиар- ный цирроз |
Пируватдегидрогеназный комплекс (РДС-Е2) |
Escherichia coli, белок РДС-Е2 |
|
Ревматическая лихорадка с вовлечением сердца |
Белки кардиального миозина |
Бета-гемолитический стрептокок, М-белок |
|
Болезнь Chagas ' |
Тяжелые цепи кардиального миозина |
Trypanosoma cruzi, белок В13 |
Однако следует учитывать, что далеко не всегда аутоиммунный ответ (особенно в виде продукции гуморальных аутоантител после инфекционных заболеваний) заканчивается развитием аутоиммунного заболевания.
Б. Согласно второму варианту теории молекулярной мимикрии, собственные (ауто-, self-) антигены хозяина могут модифицироваться под влиянием различных факторов: длительного воздействия инфекционный агентов, влияния свободных радикалов, N 0, ксенобиотиков, лекарственных средств, воздействия факторов окружающей среды (ионизирующее и ультрафиолетовое излучение, воздействие низких температур и т. п.). В результате таких воздействий аутоантигены изменяются и распознаются иммунной системой как чужеродные (nonself)- Продуцирующиеся аутоантитела и цитотоксические лимфоциты связываются не только с модифицированными аутоантигенами, но и с истинными аутоантигенами за счет все той же перекрестной реактивности (мимикрии, подобия).
В иммунологических механизмах повреждения тканей при аутоиммунный заболеваниях принимают участие все те эффекторные механизмы, которыми иммунная система защищает организм от экзогенной интервенции -гуморальные антитела, иммунные комплексы, цитотоксические Т-лимфоциты и цитокины. В развитии патологического процесса указанные факторы могут действовать как порознь, так и совместно.
При прямом действии аутоантител на клетки и ткани организма, как правило, активируется система комплемента, которая способствует их разрушению. Возможен вариант "включения" механизма антителозависимого клеточно-опосредованного лизиса, т. е. с участием К-кле- ток. В некоторых случаях аутоантитела, направленные против функционально значимых клеточных рецепторов, стимулируют или ингибируют специализированную функцию клеток без ее разрушения.
В том случае, когда формируются циркулирующие иммунные комплексы, состоящие из аутоантигена и аутоантител, разные причины могут вызвать их оседание в микроциркуляторном русле различных органов (почка, суставы, кожа и др.) или в местах гемодинамически напряженных, с выраженным турбулентным течением (бифуркации, отхождение крупных сосудов и т. п.). В местах отложения иммунных комплексов активируется комплемент, скапливаются гранулоциты и моноциты, выделяющие различные ферменты. Все это приводит к гибели клеток "шокового" органа.
Созревание цитотоксических Т-лимфоцитов приводит к их накоплению в пораженной ткани (околососудистая инфильтрация) с после. дующим развитием киллингового эффекта, привлечением большого количества клеток воспаления.
В последние годы в развитии аутоиммунного повреждения клеток и тканей большое внимание уделяют провоспалительным цитокинам — ИЛ-1, альфа-ОНФ, гамма-ИНФ, ИЛ-2, а также включению механизмов апоптоза. Сегодня имеются доказательства того, ¦ что аутоиммунные повреждения тканей могут быть реализованы за счет механизма неспецифического связывания Fas + FasL и включения апоптоза. Обусловлено это тем, что на поверхности клеток, например, В-клеток поджелудочной железы и олигодендроцитах, под воздействием различных стимулов (прежде всего, цитокинов) появляется Fas-рецептор. Аутореактивные Т-лимфоциты, экспрессирующие FasL, могут связываться с Fas-рецептором и индуцировать апоптотичеекую смерть клеток-мишеней.
Интересны также следующие наблюдения. Считают, что конститутивная (изначальная) экспрессия FasL на поверхности клеток привилегированных органов (например, глаз, яичек) носит защитный характер, позволяя индуцировать апоптоз у Fas-положительных лимфоцитов при их попадании в соответствующие ткани. Но наличие на поверхности одной и той же клетки Fas-рецептора и Fas-лиганда может быть причиной аутокринного суицида такой клетки. Подобный механизм рассматривается как одна из причин развития тиреоидита Ха- шимото (на тироцитах есть FasL, а при определенных воздействиях на мембране тироцитов начинают сильно экспрессироваться Fas-рецепторы).
Наличие аутоантител само по себе еще не указывает на развитие заболевания. В невысоких титрах аутоантитела постоянно обнаруживаются в сыворотке крови здоровых лиц и участвуют в поддержании гомеостаза, обеспечивают выведение продуктов метаболизма, идио- типический контроль и другие физиологические процессы.
Исходя из приведенных данных, можно дать определение понятий "аутоиммунный процесс" и "аутоиммунное заболевание".
Аутоиммунный процесс (аутоиммунитет) — это форма иммунного ответа, индуцированная аутоантигенными детерминантами в условиях нормы и патологии; является одним из ¦ механизмов поддержания гомеостаза. Выраженность аутоиммунных процессов в условиях нормы незначительная.
Аутоиммунное заболевание — это патологический процесс, в патогенезе которого важную роль играют аутоантитела и/или клеточный аутоиммунный ответ.
Признаки, по которым то или иное заболевание может быть отнесено к разряду аутоиммунных, сформулированы еще Л. Витебски (1961).
- Наличие аутоантител или цитотоксических Т-лимфоцитов, направленных против антигена, ассоциированного с данным заболеванием.
- Идентификация аутоантигена, против которого направлен иммунный ответ.
- Перенос аутоиммунного процесса с помощью сыворотки, содержащей антитела или цитотоксические Т-лимфоциты.
- Возможность создания с помощью введения аутоантигена экспериментальной модели заболевания с развитием соответствующих морфологических нарушений, характерных для заболевания.
Общие принципы иммуно-лабораторной диагностики аутоиммунных заболеваний основываются на следующих признаках:
- Наличие специфических аутоантител;
- Наличие специфической клеточной сенсибилизации (выявляется с помощью реакции бласттрансформации — РБТ и теста ингибиции миграции лейкоцитов в присутствии соответствующего аутоантигена);
- Повышение уровня гамма-глобулина и/или IgG;
- Изменение количества Т-хелперов и Т-супрессоров, приводящее к повышению иммунорегуляторного индекса;
- Снижение уровня СЗ и С4 компонентов комплемента;
- Отложения иммунных комплексов в пораженных тканях (IgG, IgM, СЗ, С4 и фибрин);
- Лимфоидно-клеточная инфильтрация пораженных тканей;
- Определение HLA-фенотипа.
В табл. 18 представлены некоторые аутоиммуные заболевания, антигены, на которые развивается иммунный ответ при данном заболевании и HLA-антигены, достоверно чаще встречающиеся при данном заболевании (звездочкой отмечены антигены, причинная роль которых в развитии аутоиммунитета доказана).
Таблица 18. HLA-зависимые болезни (Lechler, 1994; Yao-Hua Song и соавторы. 1996; Menard, El-Amini, 1996)
|
Болезнь |
Антиген, на который развивается иммунный ответ |
HLA- антигены |
|
Целиакий |
Альфа-глиадин |
DR3, DR7 |
|
Синдром Гудпасчера |
Коллаген базальной мембраны клубочков почки * |
DR2 |
|
Болезнь Грейвса |
Тиротропиновый рецептор * |
DR3, DR5 |
|
Тиреоидит Хашимото |
Тироглобулин *, микросомы |
DR3, DR5 |
|
Инсулинзависимый сахарный диабет |
Декарбоксилаза глутаминовой кислоты (ЦГК-65 и ДГК-67), инсулиновый рецептор, тирозинфосфатаза 1А-2 и 1А-2р, Р-клетки поджелудочной железы * |
DR3, DR4 |
|
Рассеянный склероз |
Основный белок миелина * |
DR2, DR4 |
|
Тяжелая миастения |
Рецептор к ацетилхолину * |
DR3 |
|
Болезнь Бехтерева |
Неизвестен |
В27 |
|
Синдром Рейтера |
Неизвестен |
В27 |
|
Пернициозная анемия |
Н+/К+-АТФаза, внутренний фактор *, париетальные клетки желудка |
DR5 |
|
Нарколепсия |
Неизвестен |
DR7 |
|
Прогрессирующий системный склероз |
ДНК-топоизомераза, РНК-полимераза |
DR5 |
|
Псориаз вульгарный |
Неизвестен |
DR7 |
|
Ревматоидный артрит |
Fc фрагмент IgG *, коллаген, кальпас- татин |
DR7, DR21 |
|
Ювенильный ревматоидный артрит |
Fc фрагмент IgG *, коллаген |
DR5 |
|
СКВ |
Двухспиральная ДНК *, кардиолипин, факторы свертывания, Sm, RD, RNP |
DR3, DR2 |
|
Витилиго |
Тирозиназа |
DR4 |
|
Болезнь |
Антиген, на который развивается иммунный ответ |
HLA- антигены |
|
Герпетиформный дерматит |
Глиадин, ретикулин * |
DR3 |
|
Пемфигус вульгарный |
"Ре V антигенный комплекс", эпидермис, межклеточные соединения * |
DR4 , DRw6 |
|
Ревматическая лихорадка |
Миокард (стрептококк) |
¦ — |
|
Буллезный пемфигоид |
Антиген базальной мембраны кожи * |
— |
|
Увеит |
Антиген передней камеры глаза |
— |
|
Симпатическая офтальмия |
Антиген сосудистой оболочки |
— |
|
Аутоиммунная гемолитическая анемия |
Эритроциты * |
— |
|
Идиопатическая тромбо- цитопеническая пурпура |
Тромбоциты * |
— |
|
Первичный билиарный цирроз |
Митохондрии * |
— |
|
Хронический активный |
Smooth muscle *, ядерный антиген ге- |
— |
|
гепатит |
патоцита |
|
|
Неспецифический язвенный колит |
Антиген толстой кишки |
— |
|
Болезнь Крона |
Антиген кишечника (Е. Coli) |
— |
|
Синдром Шегрена |
Ядерные антигены: SS-A, SS-B * |
— |
|
Дерматомиозит |
Ядерный антиген Go-I *, антиген скелетных мышц |
— |
|
Системная склеродермия |
Ядерный антиген Scl-70 * |
— |
|
Смешанные заболевания соединительной ткани |
Ядерный антиген RNP * |
— |