
Вирусы животных, как и фаги, способны к репродукции (от англ, reproduce - воспроизводство). Во время репродукции синтез нуклеиновых кислот и белков вируса происходит разобщенно, в разных частях клетки, затем следует сборка вирионов. Такой способ репродукции получил название дизъюнктивного.
В репродукции вирусов животных, как и бактерий, четко выделяется ряд последовательных стадий.
I стадия - адсорбция. Процесс начинается с того, что между участками адсорбции (рецепторами) вириона и рецепторами, расположенными на поверхности клетки, образуются нековалентные связи. В результате вирион адсорбируется на поверхностных структурах клетки.
У разных вирусов рецепторы неодинаковы по своей природе и расположению на вирионе. Так, у аденовирусов рецепторами служат нити, расположенные на вершинах икосаэдра. У сложных вирусов, имеющих внешнюю оболочку, рецепторами являются многочисленные шипики и ворсинки, которыми усеяна поверхность оболочки.
На поверхности клеток животных и человека также имеются разные рецепторные участки. Например, рецепторную функцию для адсорбции миксовирусов выполняют мукопротеиды оболочек, для адсорбции пикорна- и арбовирусов - липопротеиды.
- стадия - проникновение вируса в клетку хозяина. В клетки животных и человека вирусы проникают разными путями. Одни из них - путем пиноцитоза (виропексиса), сущность которого состоит в том, что в месте адсорбции вируса происходит впячивание клеточной мембраны, образуется пиноцитарная вакуоль, захватывающая вирус и «переносящая» его внутрь клетки. Вирионы, имеющие внешнюю оболочку, сливаются с клеточной мембраной и так проникают в цитоплазму.
- стадия - освобождение вирусной нуклеиновой кислоты от капсида, или «раздевание» вируса. В отличие от вирусов бактерий, вирусы высших организмов целиком проникают в клетку. Но сразу же после их внедрения под действием протеолитических ферментов клетки-хозяина происходит разрушение капсида, т. е. раздевание вируса. Некоторые вирусы начинают освобождаться от капсида во время слияния вирусной и клеточной мембран, раздевание других происходит в специализированных структурах клетки - лизосомах, ядерных порах в ядерной мембране, структурах аппарата Гольджи. В результате раздевания освобождается внутренний компонент вируса, который способен вызвать инфекционный процесс.
ГУ стадия - репликация нуклеиновых кислот и синтез вирусных белков. Эти процессы имеют свои специфические особенности у ДНК- и РНК-содержащих вирусов (см. ниже).
V стадия - созревание или сборка вир иона. Созревание вириона заключается в строго упорядоченном соединении вирусных белков и их нуклеиновых кислот. Этот сложный и необратимый процесс идет путем полимеризации белковых субъединиц и их упорядоченной укладки вокруг нуклеиновой кислоты с образованием капсида. Сборка одних вирусов, таких как пикорнавирусы, поксвирусы и большинство вирусов растений, осуществляется в цитоплазме клетки; другие же, такие как адено-, папова-, герпес- и миксовирусы, формируются в ядре. У просто организованных вирусов процесс самосборки, т. е. процесс самопроизвольного соединения вирусных нуклеиновых кислот и белков, состоит в расположении упорядоченных соединений субъединиц белка вокруг нуклеиновой кислоты. Вначале формируются про- вирионы, которые в результате модификаций белков превращаются в вирионы. Сборка сложно устроенных вирусов осуществляется многоступенчато, в ней принимают участие клеточные структуры, ядерная и цитоплазматическая мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы. При выходе из клетки эти вирусы приобретают наружную оболочку - суперкапсид.
VI стадия - выход вирионов из клетки хозяина. После завершения сборки зрелые вирусные частицы покидают клетку. Этот процесс у разных вирусов осуществляется по-разному. Большинство просто устроенных вирусов (пикорна-, рео-, парво-, аденовирусы) освобождаются из клетки в результате разрушения мембраны, деструкции клетки. Сложно устроенные вирусы освобождаются путем почкования. Например, вирионы герпеса проникают в цито-плазматическую вакуоль и с ее помощью выводятся из клетки. При этом клетка некоторое время сохраняет жизнеспособность и продуцирует вирусное потомство.
ДНК-содержащие вирусы. Репликация генома ДНК-содержащих вирусов осуществляется ДНК-полимеразами как клетки- хозяина, так и вирусной частицы.
У вирусов с двухцепочечной ДНК генетическая информация реализуется тем же путем, что и у клеточных организмов:

Развитие вируса начинается с транскрипции ранних генов, ответственных за синтез ферментов («ранних» вирусных белков), необходимых для репликации вирусной ДНК.
Вирусы, репродукция которых происходит в ядре клетки, используют для транскрипции клеточную полимеразу (папово- вирусы, аденовирусы, вирусы герпеса). Другие же, как поксвирусы (вирусы оспы), иридовирусы (вирусы лягушек), репродукция которых осуществляется в цитоплазме, не могут использовать фермент, находящийся в ядре. Они содержат собственную ДНК-
полимеразу, которая в составе вириона проникает в клетку и участвует в транскрипции вирусного генома.
Образовавшаяся иРНК транслируется рибосомами клетки- хозяина в вирусоспецифические ферменты, обеспечивающие репликацию вирусной ДНК. В результате образуется «фонд» ДНК, используемый для формирования вирусных частиц. Репликация вирусной ДНК идет по обычному полуконсервативному механизму.
При репликации однонитевых ДНК вначале происходит образование двунитевых форм, которые являются промежуточными репликативными формами. В дальнейшем они служат матрицами для синтеза вирусных однонитевых ДНК.
РНК-содержащие вирусы. Репликация нуклеиновой кислоты РНК-содержащих вирусов имеет ряд особенностей, связанных с тем, что в клетках-хозяевах нет ферментов для синтеза РНК в РНК- матрице. Поэтому РНК-содержащие вирусы в целях обеспечения собственной репродукции должны нести генетическую информацию для синтеза РНК-зависимой-РНК-полимеразы (РНК-репликазы) и РНК-зависимой-ДНК-полимеразы (обратной транскриптазы).
Репродукция РНК-содержащих вирусов также начинается с синтеза матричной РНК, которая транслируется рибосомами с образованием ферментов репликации и капсидных белков. В
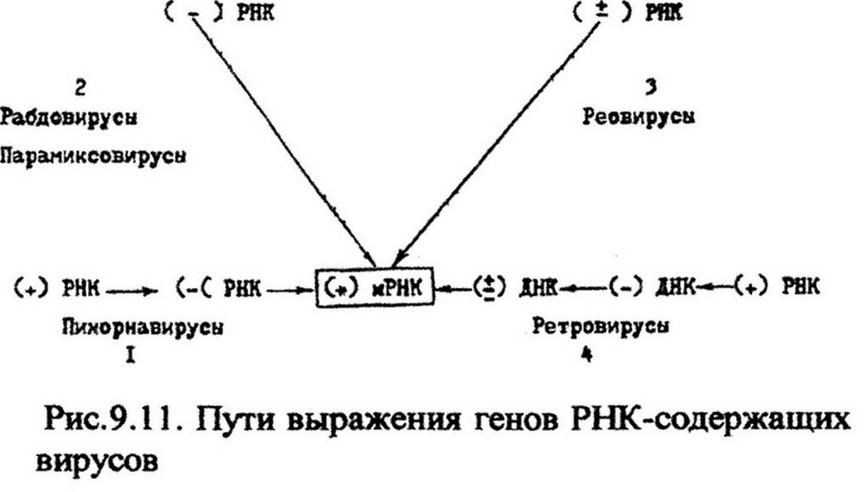 зависимости от принадлежности вируса к той или иной группе синтез мРНК может осуществляться разными путями. Известно четыре пути выражения генов РНК-содержащих вирусов (рис. 9.11).
зависимости от принадлежности вируса к той или иной группе синтез мРНК может осуществляться разными путями. Известно четыре пути выражения генов РНК-содержащих вирусов (рис. 9.11).
Пи корн а вирусы (от pico - маленький, RNA - рибонуклеиновая кислота) - группа мелких РНК-содержащих вирусов. Геном содержит одну молекулу одноцепочечной (+)РНК (плюс- цепь). Она окружена икосаэдрической белковой оболочкой.
Представителями являются вирус полиомиелита, риновирус (вызывает обычное простудное заболевание), вирус ящура. У пи- корнавирусов РНК вириона выполняет роль матрицы для синтеза вирусного белка. Попав в клетку-хозяина, она освобождается из капсида и сразу транслируется рибосомами с образованием гигантского полипептида - предшественника вирусных белков. Этот полипептид разрезается протеазами клетки-хозяина на несколько вирусоспецифических белков, среди которых имеются белки капсида и РНК-репликаза. Этот фермент синтезирует (-) цепи РНК на матрице (+)РНК вириона. (-)РНК в свою очередь служит матрицей для синтеза множества (+) цепей, которые используются в синтезе белка или укладываются в капсиды и дают новые вирионы.
Рабдовирусы (от rhabdo -палочка) - группа крупных вирусов, напоминающих по форме пулю. Геном представлен одной молекулой одноцепочечной (-)РНК, которая уложена в спиральный капсид. Капсид покрыт двуслойной липидной мембраной. Представителями являются вирус везикулярного стоматита и вирус бешенства. В составе вириона содержится фермент РНК-репликаза. Одноцепочечная (-)РНК вириона не может служить матрицей для синтеза вирусных белков. Поэтому первым этапом экспрессии генов (-)РНК является синтез (+)РНК. Этот процесс осуществляет вирусная РНК-репликаза. В случае вируса везикулярного стоматита с геномной РНК вируса транскрибируется пять моноцистронных мРНК (моноцистрон - последовательность в ДНК, кодирующая один полипептид), которые транслируются рибосомами с образованием пяти вирусных белков, входящих в состав капсида, шипов, мембраны. Вирионная РНК-репликаза синтезирует также и длинную цепь (+)РНК, содержащую всю генетическую информацию вируса. Эта (+)РНК в свою очередь служит матрицей для синтеза (-)РНК, которые используются для формирования новых вирусных частиц.
Рабдовирусы входят в группу вирусов с негативным геномом. Такое название они получили за то, что РНК вирионов даже с высокой молекулярной массой не обладает инфекционностью. Это обусловлено тем, что рабдовирусы содержат минус-цепь РНК, которая в отсутствие РНК-репликазы не способна к репликации. Как отмечалось выше, этот фермент содержится в вирионе. Для
проявления инфекционности необходимо инфицирование клетки- хозяина целостным вир ионом.
Реовирусы (от первых букв англ, слов respiratory enterie orphan, что означает энтеро-респираторный вирус-сирота, т. е. «не пристроенный» ни к какой болезни) - крупные, часто встречающиеся вирусы, но явных болезней не вызывают. Геном реовирусов состоит из 10 различных молекул двухцепочечных (+)РНК, заключенных в икосаэдрический капсид, не содержащий оболочки. РНК генома ассоциирована с белками. В вир ионе также содержится РНК-репликаза. Проникнув в клетку-хозяина, вирион освобождается от капсида. Это активирует РНК-репликазу и она начинает транскрибировать все 10 молекул РНК в (+)мРНК). Транскрипция идет асимметрично - только с образованием (-i-)mPHK («минусовые» цепи не образуются). Исходные цепи (±)РНК разрушаются. Каждая из 10 вновь образованных мРНК дает при трансляции один белок. В итоге образуется 10 разных белков. Затем все 10 мРНК соединяются с некоторыми вирусными белками и образуют предшественник сердцевины вириона, в котором синтезируются комплементарные матричные РНК минус-цепи, в результате чего формируются двухцепочечные молекулы РНК вирусов этой группы.
Ретровирусы (от англ, reverse transcriptase - обратная транскриптаза) - РНК-содержащие опухолеродные вирусы. Это единственная группа из числа РНК-содержащих вирусов, представители которой способны вызывать рак. Известными представителями ретровирусов является вирус саркомы Рауса, открытый П. Раусом в 1911 г. при изучении этиологии опухоли соединительной ткани кур. Он показал, что приготовленный из опухоли бесклеточный экстракт, пропущенный через бактериальный фильтр, при введении здоровым цыплятам вызывает у них образование такой же опухоли. Было очевидно, что опухоль вызывается вирусом.
Геном ретровирусов представлен двумя одинаковыми молекулами одноцепочечной (+)РНК, которые заключены в капсид икосаэдрической фермы. Капсид окружен оболочкой, состоящей из молекул гликопротеина, кодируемого вирусом, и двуслойной липидной мембраны, происходящей из мембраны клетки-хозяина. В вирионе содержится специфический фермент, необходимый для репродукции вируса - обратная транскриптаза, открытая Теминым и Балтимором (независимо друг от .друга) в 1970 г. Особенность этого фермента состоит в том, что он осуществляет обратную транскрипцию РНК —gt; ДНК, т. е. синтезирует двуспиральную ДНК, используя в качестве матрицы (+)РНК вириона.
После проникновения вириона в клетку вирусная (+)РНК освобождается от оболочки. Затем обратная транскриптаза использует вир ионную РНК как матрицу и синтезирует (-)цепь ДНК. Образуется гибридная молекула ДНК: одна цепь - РНК вириона, другая - вновь синтезированная цепь ДНК. Этот же фермент расщепляет цепь вирионной РНК в гибридной молекуле РНК-ДНК (рис.9.12). После этого обратная транскриптаза синтезирует вторую комплементарную цепь провирусной ДНК, матрицей служит оставшаяся первая цепь ДНК. Таким образом, обратная транск-
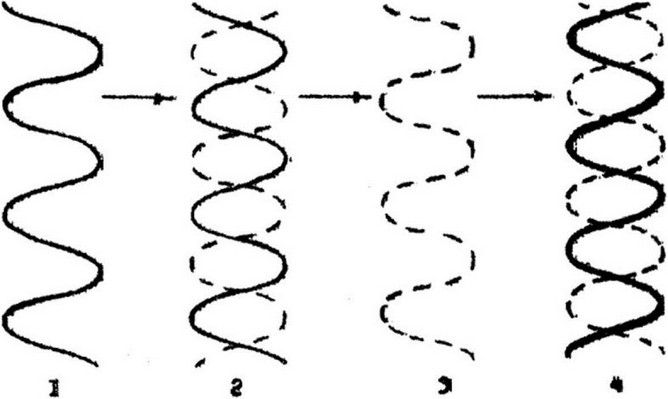
Рис.9.12. Синтез ДНК по РНК-матрице с помощью обратной транскриптазы: / - вирусная РНК; 2 - гибрид ДНК-РНК; 3 - ДНК-транскрипт вирусной РНК; 4 -
двухцепочечная вирусная ДНК
риптаза осуществляет три последовательные реакции: РНК-
зависимый синтез ДНК, гидролиз РНК и ДНК-зависимый синтез ДНК. В результате образуется ДНК-содержащий провирус, который является промежуточной формой в репликации онкогенных вирусов. Образовавшаяся двуспиральная провирусная ДНК переходит в кольцевую форму, проникает в ядро клетки и интегрирует с хромосомной ДНК. После интеграции начинается транскрипция ДНК-провируса, которая осуществляется РНК-полимеразой, ответственной за синтез мРНК в клетке. Первичным транскриптом является 35S РНК, считываемая с минус-цепи ДНК-провируса. Эта же РНК является и матрицей для синтеза вир ионных белков.
Формирование вирионов происходит на клеточной мембране путем почкования. При этом, геномная РНК и вирусные белки перемещаются к плазматической мембране и включаются в нее. Затем часть измененной мембраны отпочковывается и образует новые вирусные частицы, которые могут инфицировать новые клетки. В отличие от инфекции онкогенными ДНК-содержащими вирусами продуктивная ретровирусная инфекция не является литической. Ретровирусы обычно не убивают клеток-хозяев. Их ДНК остается в геноме зараженной клетки, интегрированная - реплицируется вместе с клеточной ДНК, поэтому дочерние клетки наследуют вирусный геном. Такие вирусы называются эндогенными. Вирусная инфекция на уровне клетки и на уровне популяции носит хронический характер. При этом, зараженные клетки могут подвергаться морфологическим и физиологическим изменениям, модифицироваться, т. е. приобретать ряд новых наследственных свойств, в том числе и способность к неограниченному росту. Модифицированные клетки образуют очаги активно делящихся клеток, которые либо превращаются в опухолевые, либо являются стадией на пути превращения нормальной клетки в опухолевую. Но только немногие штаммы онкогенных вирусов способны вызывать трансформацию клеток. Большинство из них являются дефектными - несут повреждения в одном из генов генома.
Геном РНК-содержащих опухолевых вирусов, в том числе и вируса саркомы Рауса, содержит четыре гена.
Ген gag кодирует синтез внутренних белков - группоспецифических антигенов, ген pol кодирует обратную транскриптазу, ген env кодирует гликопротеиды оболочки, необходимые для прикрепления вируса к поверхности клетки-хозяина, ген src (онкоген) - отвечает исключительно за трансформацию клетки. Мутанты, лишенные гена src (от англ, sarcoma), могут нормально размножаться, но не способны вызвать трансфермацию. В то же
время, используя лишь один ген src, можно вызвать трансформацию клеток и образование опухоли, т. е. большая часть вирусного генома в этом не принимает участия.
РНК-содержащие вирусы обладают более сильным трансформирующим действием, чем ДНК-содержащие, а в пределах группы РНК-содержащих ретровирусов - наиболее эффективны вирусы сарком. У них широкий круг хозяев и они трансформируют множество типов клеток (фибробласты, миобласты, эпителий радужной оболочки и другие).
Механизм действия продуктов опухолевых генов онкогенных вирусов еще не известен. Установлено, что продукт гена src вируса саркомы П. Рауса является протеинкиназой - ферментом, который, в отличие от других киназ, фосфор ил ирует белки не по треонину и серину, а по тирозину. Это одна из известных особенностей продукта, кодируемого геном src. Насколько значительна эта особенность в образовании опухолей - пока неизвестно.
К ретровирусам относится и возбудитель СПИДа - вирус иммунодефицита человека, или ВИЧ. Геном ВИЧ представлен РНК, причем, в зараженных клетках выявлено три класса вирусспеци- фических РНК с разной молекулярной массой.
Репликация РНК проходит через стадию образования ДНК- транскрипта, который интегрируется с геномом клетки. ВИЧ хорошо репродуцируется в Т-лимфоцитах человека.