
Вирусы бактерий носят название бактериофагов или просто фагов. Впервые явление бактериофагии в 1915 г. описал Туорт у стафилококков. Независимо от него, в 1917 г. Д’Эрелль сообщил об открытии литического агента в культуре дизентерийной палочки, выделенной от больных дизентерией людей при их выздоровлении. Он изучил биологические особенности этого агента, его взаимоотношение с бактериями и дал ему название бактериофаг, что означает «пожиратель бактерий».
Сразу после открытия бактериофагов и установления их высокой литической способности ученые занялись разработкой вопросов практического характера: использование бактериофагов для лечения и профилактики инфекционных заболеваний. Но терапевтический эффект применения бактериофагов не всегда был положительным, и с открытием антибиотиков интерес к ним как к лекарственным препаратам ослаб. Позже фаги заняли ведущее место в решении важных общебиологических проблем. Простота структурной организации фагов, высокий выход потомства в короткий промежуток времени, доступность работы с ними делают фаг весьма удобной моделью для изучения разнообразных вопросов молекулярной биологии. Они стали основным объектом генетических исследований, в первую очередь в области молекулярной генетики. На модели фага проведены классические исследование по изучению тонкой структуры гена, расшифровке наследственного кода, изучению механизма передачи наследственной информации, молекулярные основы мутационных процессов.
- Структура бактериофагов
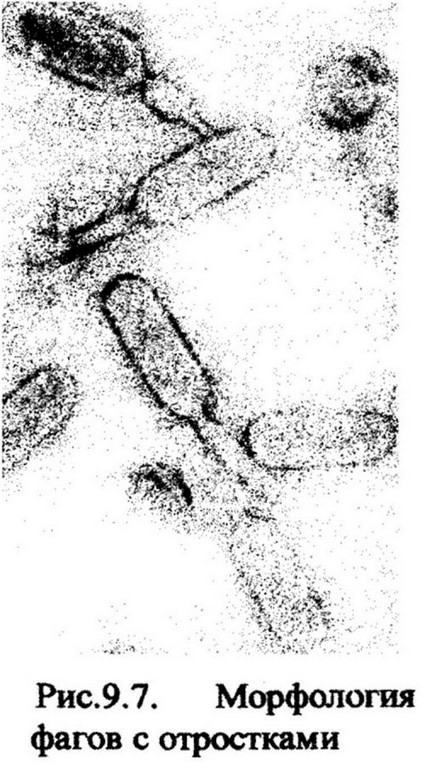 Изучение морфологии фагов началось с введения в практику биологических исследований электронного микроскопа. В 1941 г. Г. Руска впервые показал, что бактериофаги представляют собой частицы определенной
Изучение морфологии фагов началось с введения в практику биологических исследований электронного микроскопа. В 1941 г. Г. Руска впервые показал, что бактериофаги представляют собой частицы определенной
формы (рис. 9.7). Дальнейшее изучение фагов позволило установить, что они более разнообразны по форме, чем вирусы животных и растений. Среди них есть фаги нитевидной формы с коротким и длинным отростками, с аналогами отростка и др. Размеры фагов от 20 до 200 нм.
Наиболее изученными являются фаги кишечной палочки, так называемые
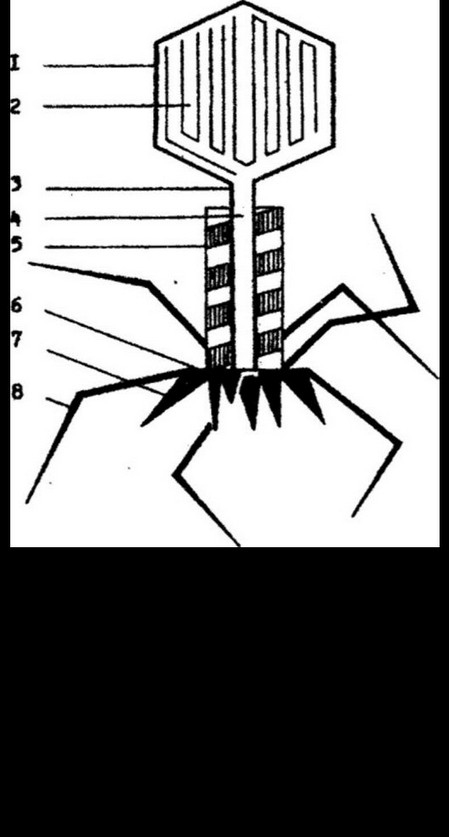 Т-фаги, которые разделяются на четные (Т2, Т4, Т6) и нечетные (Ть Т3, Т5) фаги. Наиболее сложное строение наблюдается у Т-четных фагов. У них различают головку икосаэдрической формы и отросток (рис. 9.8). Отросток представляет собой сложную структуру, состоящую из наружного чехла, внутри которого проходит тонкая полая трубка стержень. Чехол обладает способностью сокращаться. Отросток фага выполняет функцию канала, проводящего ДНК в бактериальную
Т-фаги, которые разделяются на четные (Т2, Т4, Т6) и нечетные (Ть Т3, Т5) фаги. Наиболее сложное строение наблюдается у Т-четных фагов. У них различают головку икосаэдрической формы и отросток (рис. 9.8). Отросток представляет собой сложную структуру, состоящую из наружного чехла, внутри которого проходит тонкая полая трубка стержень. Чехол обладает способностью сокращаться. Отросток фага выполняет функцию канала, проводящего ДНК в бактериальную
клетку. Длина отростка фага Т2 около ПО нм. Отросток заканчивается базальной пластинкой, несущей выросты в виде шипов. От них отходят тонкие длинные нити. Базальная пластинка и нити участвуют в процессе адсорбции и прикрепления фага на бактериальной клетке.
Основными химическими компонентами фагов являются белки и нуклеиновые кислоты. Нуклеиновые кислоты составляют содержимое головки фага. Большинство фагов включает двунитчатую ДНК, но наряду с ними существуют фаги с однонитчатой ДНК. Это мелкие сферические и нитевидные фаги. Некоторые фаги содержат РНК (фаг 7S Ps. aeruginosa, MS2, R17 Е. coli). Относительное содержание нуклеиновых кислот у фагов более высокое, чем в бактериальных клетках. Так, фаг кишечной палочки Т2 содержит 50-54 % ДНК, в то время как клетки этой бактерии - 5,2 %. Наиболее высокий процент нуклеиновой кислоты отмечается у фагов сложной структуры (30-50 %), у нитевидных фагов содержание ее
значительно меньшее (11-14 %).
Фаги различаются между собой по химическому составу ДНК. Так, в ДНК некоторых фагов выявлено наличие нестандартных оснований. В ДНК Т-четных фагов вместо цитозина содержится 5-оксиметилцитозин. В ДНК фагов Xanthomonas oryrae содержится другое производное цитозина - 5-метилцитозин. В ДНК некоторых фагов Вас. subtilis тимин заменен 5-оксиметилурацилом.
Из белков состоит оболочка головки, чехол и стержень цилиндра, базальная пластинка, нити и шипы. Кроме этих структурных белков, в отростке фага обнаружены ферментные белки. В настоящее время доказано наличие в фаговых частицах фермента лизоцима и АТФ-азы. Различают структурный и свободный фаговый лизоцим. Структурный лизоцим отличается от свободного большим молекулярным весом и функционально. Структурный лизоцим проявляет свою активность в начале фаговой инфекции - растворяет снаружи клеточную стенку бактерий, на которой адсорбировался фаг. Свободный лизоцим лизирует клетку изнутри и способствует освобождению зрелых фаговых частиц, т. е. завершает процесс фаговой инфекции.
АТФ-аза содержится в чехле фагового отростка и обеспечивает его сокращение.