
Взаимодействие вируса с клеткой, приводящее к образованию самостоятельного биологического комплекса вирус - клетка и к размножению вируса, - сложный процесс, состоящий из ряда последовательных этапов: адсорбции, внедрения, размножения, или репродукции, созревания и освобождения (рис. 9.9).
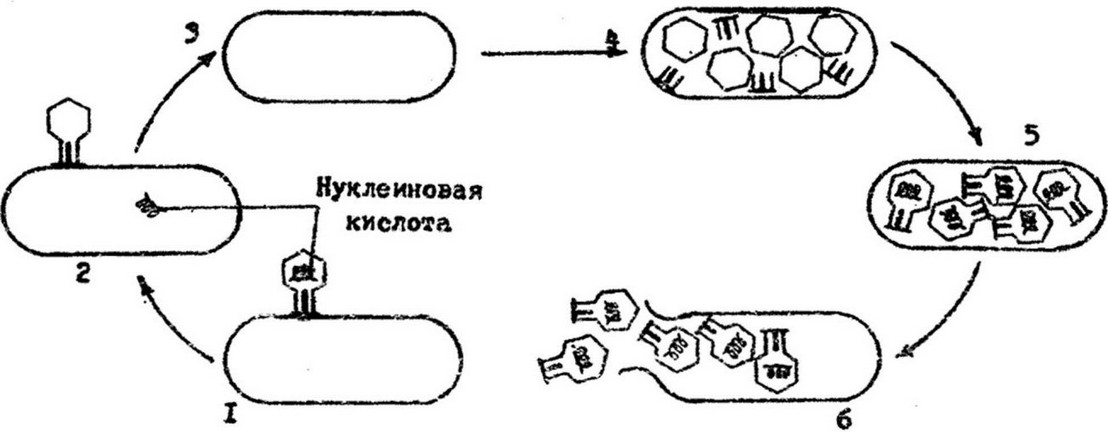
Рис.9.9. Схема взаимодействия вирулентного фага с клеткой: I - адсорбция; 2 - внедрение; 3 - начальный этап внутриклеточной репродукции; 4 - образование структурных компонентов фага; 5 - созревание фага; 6 - освобождение из клетки
У некоторых бактериофагов существует два возможных пути, по которым может пойти их развитие при заражении чувствительных бактерий: они могут размножаться и лизировать клетки - литический путь развития, или их ДНК может включаться в геном зараженной клетки, не проявляя способности к размножению и лизису, - лизогенный путь развития. Начальные этапы литического и лизогенного путей не отличаются друг от друга.
Первый этап взаимодействия фага с бактериальной клеткой - адсорбция. Он осуществляется в результате случайных столкновений фага с бактериями и прикреплении его к клеточной поверхности. Адсорбция является высокоспецифическим процессом.
Каждый вид фага адсорбируется только на определенных бактериях и на определенных участках клеточной поверхности, так называемых фагочувствительных рецепторах. Это структуры, ответственные за связывание фага. Они расположены в наружных слоях клеточной стенки. Так, рецепторы для фагов Та и Тб расположены в липопротеидном слое, для фагов Т3, Т7 и Т4 - в липополисахаридном слое клеточной стенки Е. coli. Процесс адсорбции состоит из двух стадий: первая стадия неспецифическая. Она сводится к прикреплению фаговых частиц, обусловлена электростатическими силами и носит обратимый характер. Фаговые частицы могут быть удалены с поверхности клетки при перенесении ее в среду, неблагоприятствующую адсорбции, или при обработке ее антифаговой сывороткой. Вторая стадия - специфическая, необратимая - обусловлена образованием связей между рецепторами фага, расположенными на поверхности отростка, и соответствующими рецепторами клетки. Рецепторы фага, так же как к рецепторы клетки, - это определенные химические группировки поверхностных структур. В связи с наличием множества рецепторов па одной клетке может адсорбироваться большое количестве фагов (например, ш Е. colt - до 300 фаговых частиц). На процесс адсорбции оказывают влияние как условия среды, особенно ее солевой состав, pH, так и физиологическое состояние клетки, Б период интенсивного роста бактерий способность адсорбировать фаг увеличивается. За адсорбцией фага следует этап внедрения (инъекции) фаговой нуклеиновой кислоты в клетку. Процесс начинается с сокращения чехла отростка. Сокращение стимулируется базальной пластинкой, изменяющей свою конформацию под влиянием нитей отростка. При этом лизоцим, расположенный в области базальной пластинки, разрушает' муреин клеточной стенки и внутренний стержень хвостового отростка проходит через разрыхленную клеточную стенку. Когда дистальный конец его достигает цитоплазматической мембраны, ДНК фага по каналу стержня впрыскивается в бактериальную клетку. Белковые пустые оболочки (тени фага) отрываются от клеточной стенки и разрушаются.
Третий этап - внутриклеточное размножение, или репродукция фага - заключается в синтезе компонентов вируса:
нуклеиновой кислоты и белков. Синтезируются они неодновременно, а раздельно в разных участках клетки, затем следует самосборка фаговых частиц.
С внедрением в клетку фаговой ДНК происходит перестройка метаболизма клетки в направлении синтеза компонентов фаговых частиц. Сразу после инъекции фаговой ДНК прекращается синтез бактериальных ДНК, РНК и белка, начинается синтез фагов нуклеиновой кислоты и белков. При этом. РНК-полимераза клетки транскрибирует гены фаговой ДНК в информационную РНК фага. Она служит матрицей для синтеза «ранних» белков - фагоспецифических ферментов.
Крупные и сложно организованные после внедрения в клетку фаги индуцируют синтез по крайней мере грех типов ферментов: первый тип - это нуклеазы, функция их состоит в разрушении ДНК клетки-хозяина; ферменты второго типа катализируют образование предшественников синтеза нуклеиновых кислот фага. Третий тип ферментов - это ДНК-полимеразы, РНК-репликазы и транскриптазы. Они катализируют реакции, осуществляющие репликацию и экспрессию фагового генома. ДНК фага обнаруживается в клетке через 8-10 мин после заражения. Б это время начинают синтезироваться структурные белки фага.
Четвертый этап - созревание. В этот период происходит сборка фаговой частицы, соединение ДНК фага с белковой оболочкой. Созревание начинается с уплотнения молекулы ДНК, ее конденсации и укладки. Вскоре на поверхности этой конденсированной ДНК начинают собираться молекулы субъединиц (капсомер) белковой оболочки фага и образуется капсид. К сформированной головке присоединяются отросток и его компоненты. Так образуется зрелая частица фага.
Отрезок времени с момента проникновения фаговой ДНК в бактериальную клетку и до полного созревания в ней частиц фага называется латентным, или скрытым, периодом. Каждая система бактерия-фаг имеет свой определенный латентный период. Для коли-дизентерийных фагов он составляет 15-20 мин, для фагов микобактерий - до 75 мин. В конце латентного периода под действием свободного лизоцима происходит растворение клеточной
стенки бактерий изнутри, клетка разрывается и потомство фага высвобождается в окружающую среду.
Фаги, которые в процессе взаимодействия с клеткой вызывают ее гибель, называются вирулентными. Наряду с ними существует другой тип фагов. Они также заражают бактериальную клетку, но не размножаются в ней автономно и не лизируют ее. Эти фаги называются умеренными или симбиотическими. В их взаимоотношении с клеткой отмечаются только первые два этапа: адсорбция и внедрение. После внедрения нуклеиновая кислота умеренного фага встраивается в хромосому бактерии и в течение длительного времени может реплицироваться вместе с ней и передаваться потомству. Фаг в интегрированном с хромосомой состоянии называют профагом.
Итак, различают три состояния фага: зрелый фаг (фаг находится вне клетки и способен к заражению чувствительных бактерий), профаг (ДНК фага включена в бактериальную хромосому), вегетативный фаг (фаг находится в клетке в состоянии размножения).
Бактериальные клетки, содержащие профаг, называются лизогенными, а процесс включения генома фага в геном бактерии - лизогенизацией, само же явление - лизогенией. Лизогения рассматривается как наследственное свойство бактерий продуцировать фаг. Иногда профаг спонтанно с частотой 10-10 может перейти в вегетативный фаг, т. е. происходит его размножение по типу вирулентного фага. В результате клетка лизируется и частицы фага высвобождаются и заражают новые бактерии.
Особенностью лизогенных бактерий является их иммунитет к заражению тем фагом, по которому они лизогенны, т. е. который содержится в них в состоянии профага. В основе иммунитета лежит образование в цитоплазме клетки особого вещества белковой природы - репрессора, препятствующего превращению профага в вегетативное состояние. Репрессор ингибирует репликацию и созревание фага. Синтез репрессора контролируется генами профага. Однако действие репрессора может быть снято путем обработки лизогенных бактерий специфическими агентами, которые либо взаимодействуют с нуклеиновыми кислотами, в том числе и с генетической областью профага, контролирующей синтез репрессора, либо воздействуют непосредственно на молекулы репрессора и инактивируют их. И в том и в другом случае продукция фага лизогенными бактериями увеличивается. Данное явление называют индукцией фага, а агенты - индуцирующими. К индуцирующим агентам относятся ультрафиолетовые лучи (УФ), алкилирующие соединения, температура и др.
Индукция лизогенной культуры с образованием фагового потомства может происходить при конъюгации, если донорская клетка лизогенная, а реципиентная -нелизогенная. Это значит, что в реципиентной клетке отсутствует репрессор репликации фаговой ДНК. Поэтому сразу после переноса профага в составе хромосомы в реципиентную клетку и образования мерозиготы начинается индукция перенесенного профага в фаг и образование фагового потомства. Это явление получило название зиготной индукции.
Способность умеренных фагов к лизогенизации определяется способностью индуцировать синтез собственного репрессора.
Многие лизогенные культуры содержат по 2-4 умеренных фага, т. е. являются полилизогенными. Наличие таких культур известно среди актиномицетов, клубеньковых бактерий, некоторых споровых бактерий. В связи с тем, что при лизогенизации умеренный фаг, точнее его нуклеиновая кислота, включается в генетический аппарат клетки, в последней могут наблюдаться изменения некоторых свойств. Например, при лизогенизации фагом р палочка дифтерии приобретает способность образовывать токсин. Токсичность сальмонелл также связана с наличием в клетках определенных фагов. Изменения свойств бактерий, происходящие в результате лизогенизации клеток, получили название лизогенной конверсии. Эти изменения наблюдаются только до тех пор, пока в клетке есть профаг. Если клетка теряет профаг, она теряет и приобретенные свойства.