
Зрелая частица вирусов носит название вирион. У просто организованных вирусов вирион включает два компонента: нуклеиновую кислоту и белок. Нуклеиновая кислота составляет внутреннее содержимое вириона, а белок образует наружную оболочку - капсид (от греч. capsa - футляр). Вирионы сложно организованных вирусов на поверхности капсида имеют дополнительную внешнюю оболочку - суперкапсид липопротеид- ной природы.
Капсид вирионов состоит из белковых субъединиц - капсомеров. Число их в капсидах различных вирусов неодинаковое, но постоянное для вирусов одной и той же группы. Оно определяется общим количеством белковых субъединиц, входящих в состав капсида, и характером капсомеров. Например, при наличии в капсиде 60 белковых субъединиц, где каждый капсомер состоит из 4 субъединиц, т. е. тетрамеров, число капсомеров будет равно 15 (60 : 4). Сложные изометрические капсиды (капсиды, имеющие форму многогранника, например, икосаэдра) как правило образованы капсомерами двух типов - пентамерами, расположенными в вершинах икосаэдра, и гексамерами, заполняющими его треугольные грани. Так, капсид аденовирусов имеет 252 капсомера, из которых 12 пентамеров и 240 гексамеров. Размер капсида определяет размер вириона. Например, капсид вируса полиомиелита
имеет 60 капсомеров, вируса герпеса - 162 капсомера, вируса табачной мозаики - около 2000 капсомеров.
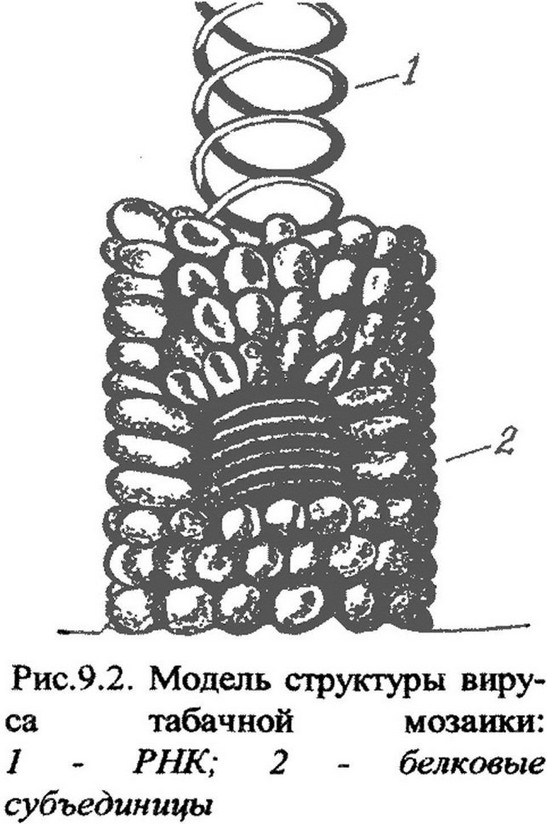 Вирионы большинства вирусов характеризуются симметричной укладкой капсомеров в капсиде. Особенность укладки капсомеров определяет тип симметрии. Он может быть спиральный, кубический или смешанный. При спиральном типе капсомеры располагаются в виде спирали вокруг винтообразно уложенной нуклеиновой кислоты (рис. 9.2). Такой тип симметрии имеют цилиндрические и нитевидные вирусы. При кубическом типе симметрии капсомеры располагаются по граням капсида и приобретают форму икосаэдра (двадцатигранника). Этот тип симметрии имеют вирусы полиомиелита, ящура, некроза табака. Смешанный тип симметрии характерен для сложных вирусов: гриппа, кори, некоторых фагов.
Вирионы большинства вирусов характеризуются симметричной укладкой капсомеров в капсиде. Особенность укладки капсомеров определяет тип симметрии. Он может быть спиральный, кубический или смешанный. При спиральном типе капсомеры располагаются в виде спирали вокруг винтообразно уложенной нуклеиновой кислоты (рис. 9.2). Такой тип симметрии имеют цилиндрические и нитевидные вирусы. При кубическом типе симметрии капсомеры располагаются по граням капсида и приобретают форму икосаэдра (двадцатигранника). Этот тип симметрии имеют вирусы полиомиелита, ящура, некроза табака. Смешанный тип симметрии характерен для сложных вирусов: гриппа, кори, некоторых фагов.
Капсид вирионов выполняет защитную функцию - предохраняет нуклеиновую кислоту вируса от действия нуклеаз и физических факторов, а также определяет хемосорбцию вируса на поверхность клетки хозяина. Эту функцию он выполняет благодаря наличию в его составе рецепторов, комплементарных рецепторам заражаемой клетки.
Химический состав вирусов несложный. Большинство из них включают только нуклеиновую кислоту и белок. Как уже отмечалось, вирусы содержат лишь один тип нуклеиновой кислоты - ДНК или РНК. И тот и другой тип является носителем наследственной информации, выполняя таким образом функции генома. Вирусы отличаются от клеточных организмов наличием уникальных форм нуклеиновых кислот - двуспиральной РНК (вирусы некоторых опухолей растений) и односпиральной ДНК
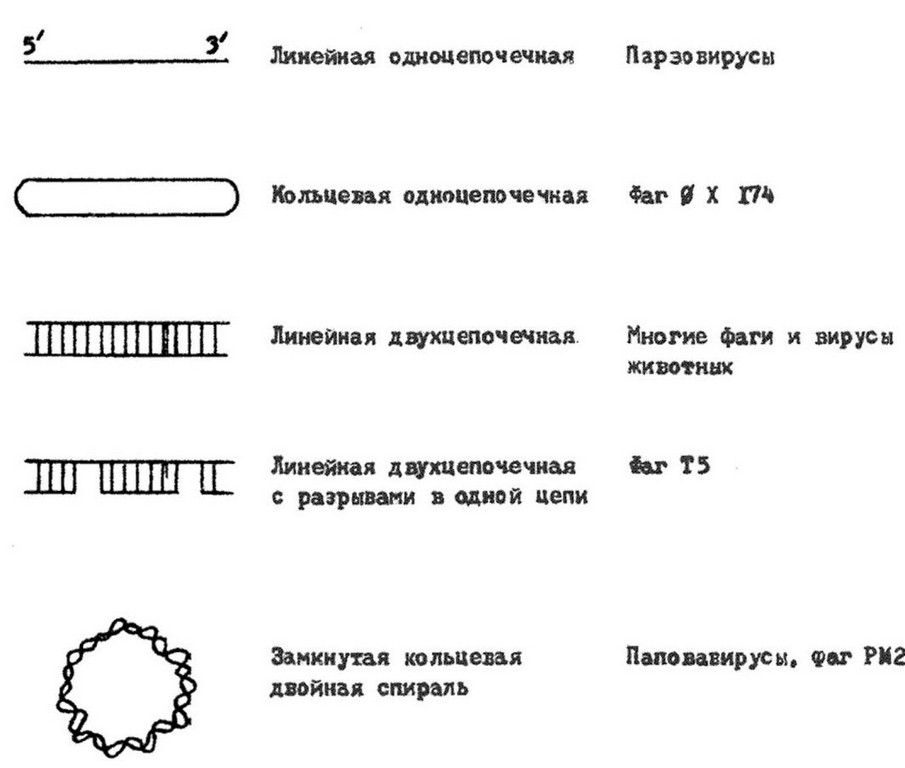
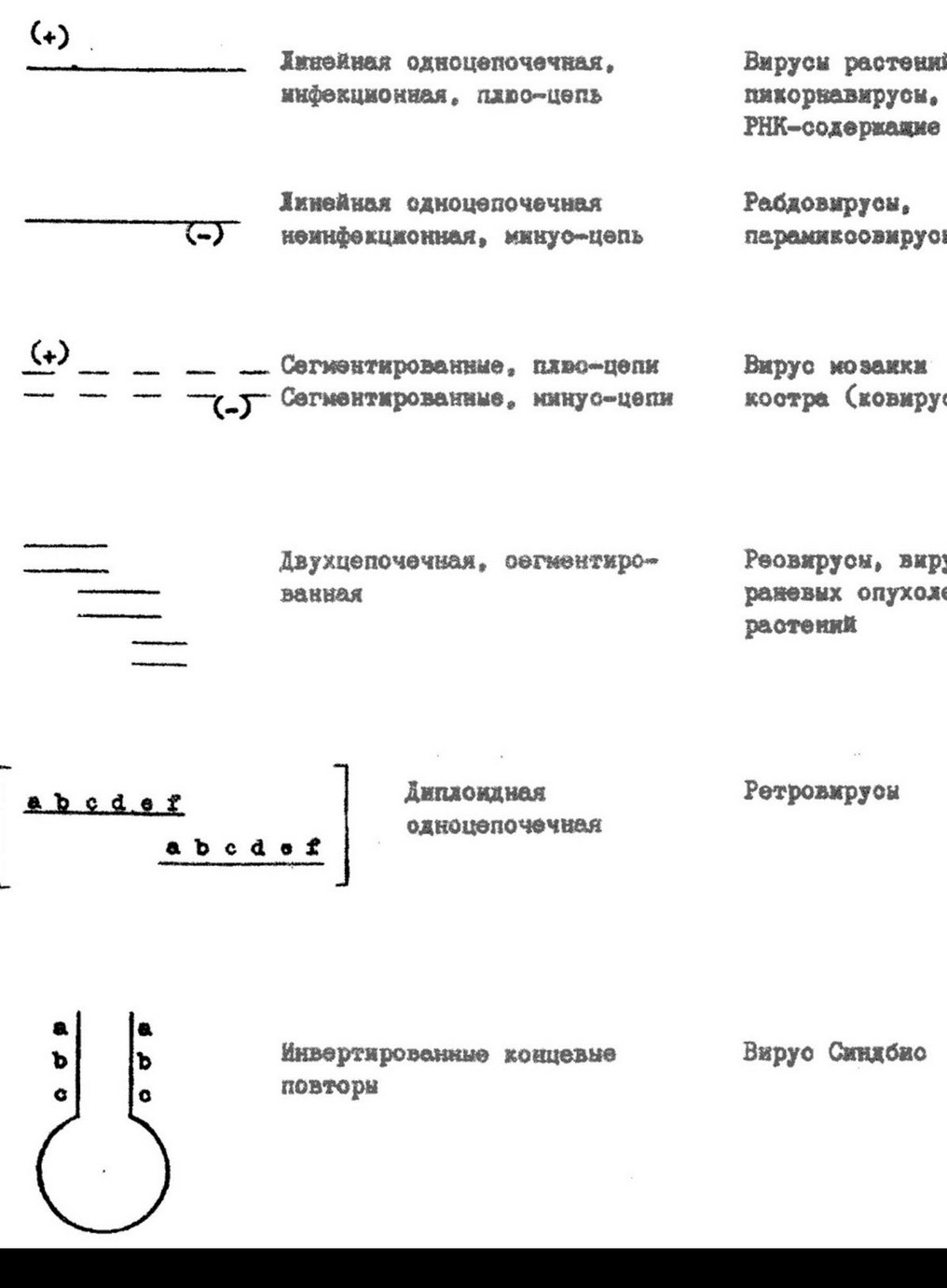
(мелкие и нитчатые фаги). Нуклеиновым кислотам вирусов присущ ряд специфических особенностей. Молекулы их могут быть линейными и кольцевыми, непрерывными и фрагментированными (рис. 9.3, 9.4). Так, вирионы всех РНК-содержащих вирусов и большинства ДНК-содержащих обладают линейными молекулами нуклеиновой кислоты. Кольцевую форму ДНК имеют вирусы папилломы, полиомы фаг 0X174. У данного фага молекула ДНК линейна, но после проникновения в клетку она замыкается в кольцо. Это обусловлено наличием «липких» (комплементарных) одноцепочечных концов, которые воссоединяются, и ДНК замыкается в кольцо. Образование кольца предшествует интеграции ДНК фага в геном клетки.
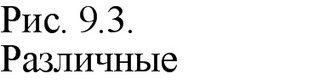
*•
 фата
фата
i fOU
ДНК ряда вирусов (фаг РМ2, вирус мозаики цветной капусты) способна к суперспирализации, что обеспечивает упаковку значительного количества
ДНК вирусов характеризуется большим разнообразием концевых участков (рис.9.5). Это различные типы избыточной последовательности нуклеотидов, концевые повторы (повторение в конце молекулы нуклеотидов ее начального фрагмента) прямых и инвертированных (перевернутых в обратном направлении). Различия в повторяющихся последовательностях нуклеотидов на концах молекул ДНК играют определенную роль в инициации транскрипции и репликации ДНК.

Рис. 9.5. Типы кольцевых участков вирусной ДНК
ДНК вирусов, в частности бактериофагов, содержит заметные количества нетипичных оснований. Например, ДНК Т-четных фагов вместо цитозина содержит 5’- оксиметилцитозин, часто глико- зил ирован. Присоединение остатка глюкозы к гидрокси-
метилцитозину препятствует разрушению ДНК фага ферментами клетки хозяина. Метилирование фаговой ДНК также защищает ее от действия бактериальных рестриктаз, которые расщепляют чужеродную ДНК. Существуют большие различия в размерах вирусных ДНК. Так, молекулярная масса ДНК фага кодирует только 9 белков.
У Т-четных фагов, вирусов осповакцины и герпеса масса ДНК до
8
2,5 * 10 дальтон. Она может кодировать более 100 белков (у фага Т4 уже идентифицировано более 100 генов).
У некоторых РНК-содержащих вирусов выявлены сегментированные одноцепочечные и двухцепочечные молекулы РНК (см. рис. 9.4). Причем, у одних вирусов все фрагменты РНК находятся в одной частице (некоторые реовирусы, миксовирусы, ретровирусы), у других - в разных частицах. Такие вирусы, нуклеиновая кислота которых «распределена» по нескольким частицам, называются ковирусами (комплементарные многокомпонентные вирусы). Представителями являются вирус мозаики коровьего гороха, мозаики люцерны, погремковости табака и другие. Например, РНК вируса мозаики коровьего гороха состоит из двух фрагментов, каждый из них находится в разных частицах. По отдельности ни одна частица не обладает инфекционностью, так как из-за отсутствия полного генома не способна к репродукции. Информация, необходимая для образования полноценных вибрионов, распределена между молекулами РНК обеих частиц. Поэтому для репродукции вируса необходимо, чтобы в одну и ту же клетку попали обе частицы. У вируса мозаики огурца фрагменты РНК распределены в трех частицах, и для репродукции его необходимо наличие в клетке всех трех частиц.
Вирусы содержат неодинаковые в функциональном
отношении геномные одноцепочечные РНК. Одни из них могут выполнять функцию информационной РНК и служить матрицей для трансляции. Такой тип РНК получил название «плюс»-цепь. Другой тип вирусных одноцепочечных РНК, так называемые «минус»-цепи, в трансляции непосредственного участия принимать не могут. Матрицей для трансляции у таких вирусов служит комплементарная «минус»-цепи информационная РНК.
Вирусы, содержащие «минус»-цепи, при инфицировании клетки привносят в нее фермент РНК - зависимую РНК-полиме- разу, который в клетке отсутствует. Известны так называемые вирусы-сателлиты, которые отличаются крайне низким содержанием нуклеиновой кислоты, и репродукция их полностью зависит от других неродственных вирусов. Примером может служить вирус- сателлит полноценного вируса некроза табака. Это очень мелкий дефектный вирус, РНК которого содержит всего 1200 нуклеотидов, что достаточно только для кодирования белка капсида. Самостоятельно он не способен к репродукции. Размножение его происходит только в том случае, если в этой же клетке растения находится его «помощник» - вирус некроза табака. Последний индуцирует образование РНК-полимеразы и других компонентов, необходимых для репликации РНК вируса-сателлита.
В 1971-1973 гг. выявлены новые, еще более мелкие инфекционные агенты - возбудители болезней растений, названные вироидами. Это «голые», т. е. лишенные белковой оболочки, небольшие кольцевые молекулы одноцепочечной РНК с молекулярной массой 105 дальтон. Их «геном» содержит 300-400 нуклеотидов, что достаточно только для кодирования белка с молекулярной массой 10000. Однако несмотря на низкое содержание генетической информации, вироиды реплицируются в клетке самостоятельно (не нуждаются в помощниках). Репликация их происходит в ядре инфицированной клетки при помощи ее же активированных ферментов. Вироиды являются возбудителями болезней картофеля, огурцов, цитрусовых, кокосовых пальм и других растений.
Белки всех исследованных вирусов относятся к глобулинам и состоят из обычных L-аминокислот. Среди них преобладают кислые ди карбоновые аминокислоты. Особенностью вирусных белков является высокая устойчивость к действию протеолитических ферментов. Это обусловлено структурными особенностями свободных концевых амино- и карбоксильных групп в пептидных цепях. Большинство концевых аминогрупп ацетилировано, а карбоксильные находятся в замаскированном состоянии благодаря особому расположению пептидных цепей. Вирусные белки обладают высокой устойчивостью к действию физических и химических факторов. Различают две группы белков, синтез которых кодируется вирусным геномом: структурные белки - входят
в состав вирусной частицы, неструктурные белки - обслуживают процесс внутриклеточной репродукции вируса, но в состав вирусных частиц не входят. Структурные белки в свою очередь делятся на капсидные и суперкапсидные. К капсидным относятся белки, образующие капсид, а также геномные белки и ферменты, входящие в состав капсида. Геномные белки ковалентно связаны с концом вирусной нуклеиновой кислоты и участвуют в регуляции функций генома. Геномные белки выявлены в составе капсида аденовирусов и др.
У некоторых сложно устроенных вирусов в составе капсида имеются ферменты транскрипции и репликации нуклеиновых кислот - РНК- и ДНК-полимеразы, ферменты, модифицирующие концы иРНК.
Суперкапсидные белки являются гликопротеидами, располагаются в суперкапсиде, формируя «шипы» на поверхности вирусной частицы. Они выполняют функцию рецепторов, осуществляя распознавание адекватных клеточных рецепторов, необходимых для адсорбции вируса на клетке. Эти гликопротеиды получили название вирусных прикрепительных белков.
Второй функцией суперкапсидных гликопротеидов является участие в слиянии вирусной и клеточной мембран, в процессе, обеспечивающем проникновение вируса в клетку.
К неструктурным белкам относятся предшественники вирусных белков, которые отличаются от других белков нестабильностью в зараженной клетке в силу быстрого нарезания их на структурные белки. К этой же категории белков относятся ферменты синтеза нуклеиновых кислот - РНК- и ДНК-полимеразы, ферменты, модифицирующие вирусные белки, - протеиназы и протеин- киназы, белки-регуляторы. Типы структурных и неструктурных белков вирусов разной сложности организации показаны на рис. 9.6.
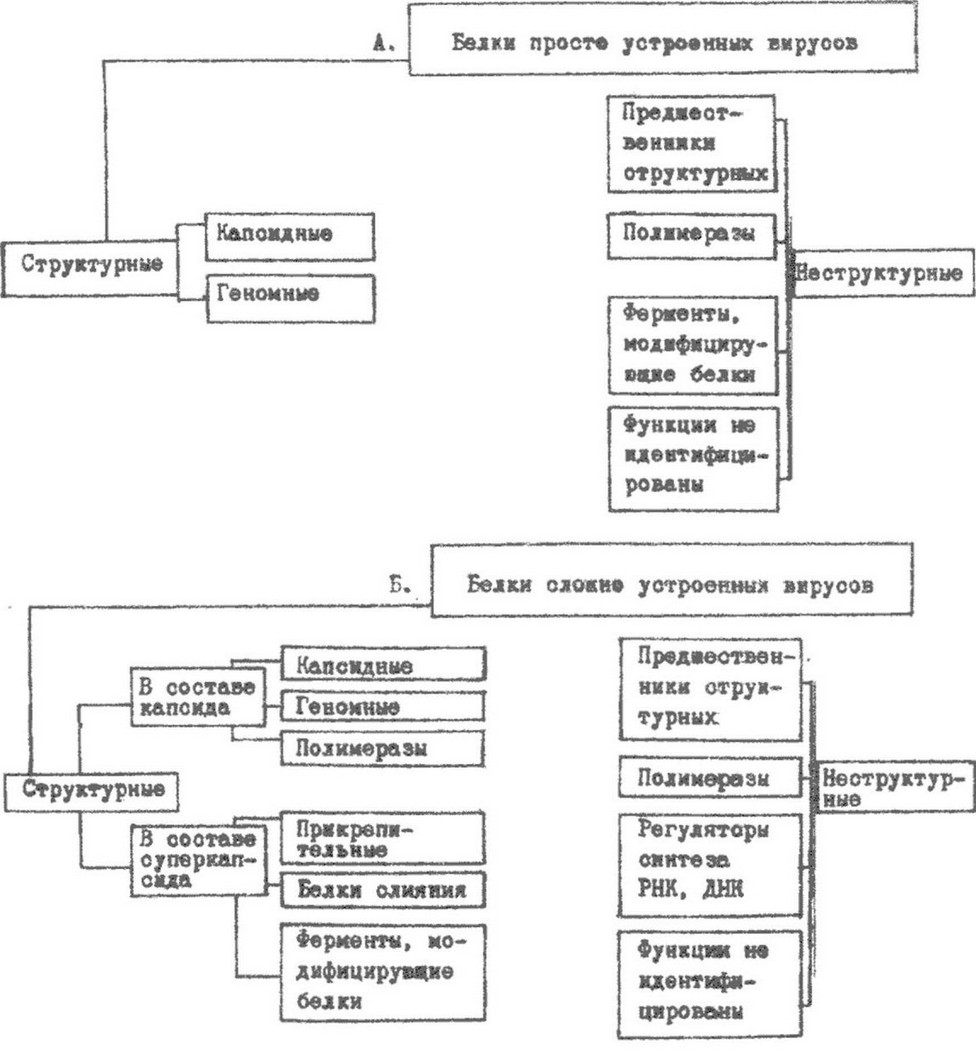
Рис. 9.6. Белки просто и сложно устроенных вирусов
Кроме белка и нуклеиновой кислоты сложно организованные вирусы содержат липиды и углеводы. Липиды входят в состав суперкапсида, формируя его липидный бислой, в который погружены суперкапсидные белки. Липиды происходят из мембран клеток-хозяев. Они стабилизируют структуру вирусной частицы. Обработка вирусной частицы липазами приводит к деградации и потере инфекционности.
Углеводы входят в состав гликопротеидов сложно организованных вирусов. Как и липиды, они имеют клеточное происхождение. Углеводный компонент гликопротеидов обеспечивает
сохранение конформации белковой молекулы, защищает ее от действия протеаз.