
Под клеточной стенкой бактерии расположена цитоплазматическая мембрана (ЦПМ). Она отделяет содержимое клетки от клеточной стенки и является обязательной структурой любой клетки.
Толщина ЦПМ бактерий обычно около 6-8 нм. На ее долю приходится до 15 % сухой массы клетки. Она состоит из липидов (15-45 %), белков (45-60 %) и небольшого количества углеводов (около 10 %). Липиды представлены фосфолипидами - до 30 % сухой массы мембраны. Среди них преобладают фосфатидил- глицерин и дифосфатид ил глицерид (кардиолипин) - обязательный компонент митохондриальных мембран эукариот. В меньшем количестве содержатся фосфатидилинозит и фосфатид ил-
этаноламин. Кроме фосфолипидов в мембране обнаружены различные гликолипиды, небольшие количества каротиноидов и хинонов. В составе липидов, производных глицерина, выявлены нетипичные для мембран жирные кислоты - насыщенные или мононенасыщенные с 16 - 18 углеродными атомами, а также кислоты, не встречающиеся в мембранах эукариот - циклопропановые и разветвленные жирные кислоты с 15-17 углеродными атомами. Набор жирных кислот, как и состоящих из них липидов мембран, является видоспецифичным для прокариот.
Мембранные липиды представлены небольшими полярными молекулами, несущими гидрофильные (головки) и гидрофобные (хвосты) группы. В водной среде они спонтанно образуют замкнутый бимолекулярный слой - бислой. Этот слой служит существенным барьером для ионов и полярных соединений. Организованные в бимолекулярный слой липиды составляют структурную основу мембраны, поддерживают механическую стабильность и придают ей гидрофобность.
Белки составляют больше половины сухой массы мембраны. Их насчитывается более 20 различных типов. Исходя из различий в прочности связи с липидами и расположением в мембране, белки подразделяют на интегральные и периферические. Интегральные белки погружены в гидрофобную область мембраны, где образуют многочисленные связи с углеводородными цепями липидов,
создавая липопротеидные комплексы. Периферические белки локализованы на поверхности гидрофильного слоя и часто присоединяются к интегральным белкам (рис. 3.14).
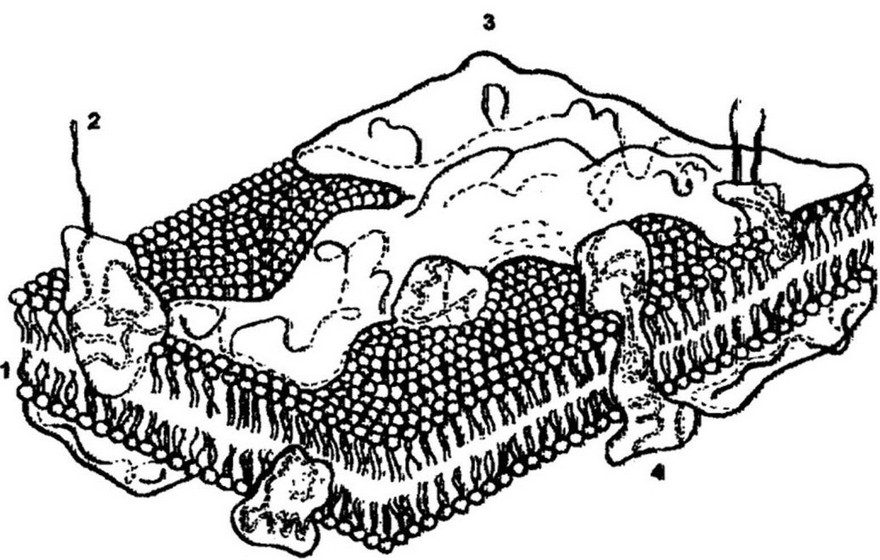
Рис.3.14. Структура цитоплазматической мембраны: 1 - липиды; 2 - гликопротеиды; 3 - периферические белки; 4 - интегральные белки
Мембранные белки по их функциям в составе мембран можно разделить на две группы: структурные и динамические.
Функции структурных белков ограничиваются поддержанием структурной целостности мембраны. Они располагаются на поверхности гидрофильного липидного слоя, выступая в роли молекулярного бандажа.
К динамическим относят белки, которые непосредственно участвуют во всех процессах, происходящих на мембране. Их разделяют на три класса: транспортные, участвующие в транспорте соединений внутрь и наружу клетки; каталитические, выполняющие функции ферментов в реакциях, происходящих на мембране; белки- рецепторы, специфически связывающие определенные соединения (токсины, гормоны) на наружной стороне мембраны.
Углеводы в мембране находятся не в свободном состоянии, а взаимосвязаны с белками и липидами в гликопротеиды. Они, как
правило, локализованы только на наружной поверхности мембраны и выполняют функции рецепторов узнавания факторов внешней среды.
Цитоплазматическая мембрана бактерий, как и все другие биологические мембраны, является асимметричной жидкокристаллической структурой. Асимметрия обусловлена различиями в химическом строении молекул белка и их расположении в липидном бислое мембраны. Одни белки расположены на поверхности бислоя, другие - погружены в его толщу, третьи проходят насквозь от внутренней до внешней поверхности бислоя. Строго определенная ориентация мембранных белков в свою очередь обусловлена тем, что они синтезируются и включаются в мембрану асимметрично. Наружная и внутренняя поверхности мембраны различаются также по ферментативной активности. В зависимости от условий (например, температуры) ЦПМ может находиться в различных фазовых состояниях: разжиженном или кристаллическом. При переходе одной жидкокристаллической фазы в другую изменяется подвижность компонентов мембраны и плотность ее упаковки, что, в свою очередь, приводит к нарушению ее функциональной активности.
Структурная организация и функции цитоплазматической мембраны. Для объяснения природы и механизма многочисленных функций ЦПМ наиболее подходящей является жидкостно-мозаичная модель организации биологических мембран, предложенная Р. Сингером и А. Николсоном в 1972 г. Согласно данной модели, мембраны представляют собой двумерные растворы определенным образом ориентированных глобулярных белков и липидов. Липиды образуют бислой, в котором гидрофильные «головки» молекул обращены наружу, а гидрофобные «хвосты» погружены в толщу мембраны, обладая при этом достаточной гибкостью. Мембранные липиды и многие белки свободно перемещаются в бислое, но только в латеральном направлении (латеральная диффузия). В поперечном направлении, т. е. от одной поверхности мембраны к противоположной, белки перемещаться не могут, а липиды перемещаются крайне медленно (1 раз за несколько часов). Причиной отсутствия или низкой активности поперечной диффузии, по-видимому, является асимметричное распределение липидов:
одних липидов больше в наружной части бислоя, других - во внутренней. Следствием этого является неодинаковая электронная плотность (проводимость) бислоя в поперечном направлении.
В жидкокристаллическом или разжиженном состоянии ЦПМ находится только при определенных, так называемых
биологических температурах. При понижении температуры ( ниже точки плавления, Тпл) липиды переходят в кристаллическое состояние, повышается степень вязкости вплоть до затвердевания мембраны. Значение температуры, вызывающей затвердевание мембраны, определяется содержанием ненасыщенных и
разветвленных жирных кислот. Чем больше их в мембране, тем ниже температура перехода липидов из жидкокристаллического состояния в кристаллическое.
Прокариоты обладают способностью регулировать текучесть мембраны путем изменения числа двойных связей и длины цепей молекул жирных кислот. Так, у Е. coli при понижении температуры среды от 42° С до 27° С соотношение насыщенных и ненасыщенных жирных кислот в мембране снижается с 1,6 до 1,0, т. е. содержание ненасыщенных жирных кислот достигает уровня насыщенных. Это предотвращает увеличение вязкости и обеспечивает сохранение клетками физиологической активности при пониженной температуре.
ЦПМ выполняет у прокариот многочисленные жизненно важные функции. В основном они определяются локализованными в ней белками, которые выполняют роль каналов, рецепторов, регенераторов энергии, ферментов, транспортные функции и другие. ЦПМ является основным осмотическим барьером, который, благодаря наличию механизмов мембранного транспорта, осуществляет избирательное поступление веществ в клетку и удаление из нее продуктов метаболизма. Избирательная проницаемость ЦПМ обусловлена локализованными в ней субстратспецифическими пермеазами, осуществляющими активный перенос через мембрану различных органических и минеральных веществ. В ЦПМ содержатся ферменты биосинтеза мембранных липидов и макромолекул, входящих в состав клеточной стенки, наружной мембраны, капсулы. ЦПМ является местом локализации окислительно-восстановительных ферментов, осуществляющих
транспорт электронов, окислительное и фотосинтетическое фосфорилирование, генерирование электрохимической энергии
трансмембранного потенциала (А// +) и химической (АТФ). ЦПМ
н
выполняет важные функции в биосинтезе и транслокации секретируемых белков грамотрицательными бактериями. Биосинтез данных белков осуществляется на рибосомах, прикрепленных к ЦПМ. У грамотрицательных бактерий на ЦПМ имеются специальные рецепторные белки, «узнающие» сигналы из большой рибо- сомной субчастицы о прикреплении рибосомы и начале синтеза белка. Мембранные рецепторные белки взаимодействуют с большой субъединицей рибосомы, образуется рибосомомембранный комплекс, на котором осуществляется синтезсекретируемых белков. Таким путем, например, Е. coli синтезирует щелочную фосфотазу, Вас. subtilis - а-амилазу. ЦПМ обеспечивает также перенос данных белков в периплазматическое пространство. Велика роль ЦПМ в регуляции клеточного деления, репликации хромосомы и плазмид и последующей сегрегации этих генетических элементов между вновь образующимися дочерними клетками.
Все прокариоты наряду с цитоплазматической мембраной содержат ее производные - внутриклеточные мембраны, которые выполняют специализированные функции. Цитоплазматическая мембрана способна к образованию всевозможных инвагинаций (впячиваний). Эти инвагинации составляют внутриклеточные мембраны, которые имеют различную протяженность, упаковку и локализацию в цитоплазме. Они могут быть собраны в сложные клубки - пластинчатые, сотовидные или трубчатые образования. Менее сложные мембраны имеют вид простых петель или канальцев различной протяженности. Независимо от сложности организации внутриклеточных мембран, все они являются производными цитоплазматической мембраны. Величина активной поверхности их превышает таковую цитоплазматической мембраны. Это дает основание судить о большой функциональной активности данных структур в клетках.
Особенно богатый внутриклеточный мембранный аппарат обнаружен у азотфиксирующих и фотосинтезирующих бактерий, бруцелл, нитрифицирующих бактерий. У фотосинтезирующих бактерий (Rhodospirillum rubrum) мембраны имеют вид замкнутых пузырьков - везикул. Их образование начинается с впячивания цитоплазматической мембраны, которое затем образует трубочку. На трубочке появляются перетяжки, разделяющие ее на ряд пузырьков. Эти пузырьки называют хроматофорами. В них содержатся поглощающие свет пигменты - бактериохлорофиллы и каротиноиды, ферменты транспорта электронов - убихиноны и цитохромы, компоненты системы фосфорилирования. У некоторых фотосинтезирующих прокариот, в частности у пурпурных серобактерий и цианобактерий, фотосинтезирующий аппарат представлен стопками мембран, которые имеют уплощенную форму и по аналогии с гранами хлоропластов зеленых растений называются тилакоидами (рис. 3.15).
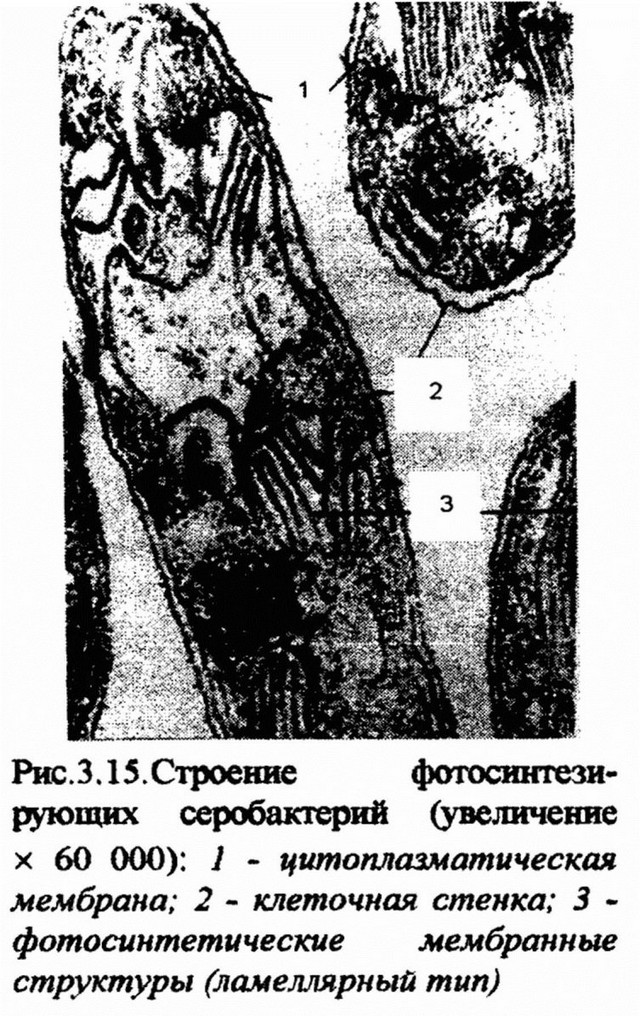 В них концентрируются пигменты фотосинтеза, ферменты электрон- транспортной цепи и системы фосфорилирования. Особенностью тила- коидов цианобактерий является отсутствие связи с цитоплазматической мембраной. Это единственная группа прокариот, имеющая дифференцированную мембранную систему.
В них концентрируются пигменты фотосинтеза, ферменты электрон- транспортной цепи и системы фосфорилирования. Особенностью тила- коидов цианобактерий является отсутствие связи с цитоплазматической мембраной. Это единственная группа прокариот, имеющая дифференцированную мембранную систему.
У нитрифицирующих бактерий внутриклеточный мембранный аппарат имеет вид пластинок, или ламелл, состоящих из плоских пузырьков (рис. 3.16).
 Из внутриклеточных мембран наиболее сложную структуру имеют мезосомы. Они представляют собой спиралевиднозакрученные, плоские или сферически трубчатые тельца. Формируются мезосомы в период клеточного деления в зоне образования поперечной перегородки. Они принимают участие в репликации хромосомы и распределении геномов между дочерними клетками, в синтезе веществ клеточной стенки. На участие
Из внутриклеточных мембран наиболее сложную структуру имеют мезосомы. Они представляют собой спиралевиднозакрученные, плоские или сферически трубчатые тельца. Формируются мезосомы в период клеточного деления в зоне образования поперечной перегородки. Они принимают участие в репликации хромосомы и распределении геномов между дочерними клетками, в синтезе веществ клеточной стенки. На участие
мезосомы в делении клетки указывает связь ее с ДНК нуклеоида. Хорошо развитые мезосомы обнаруживаются только у грам- положительных бактерий.
Накопленные к настоящему времени сведения говорят о том, что мембранные структуры бактерий достаточно дифференцированы и обеспечивают ход различных метаболических процессов в клетке.
Цитоплазма - полужидкая коллоидная масса, состоящая на 70-80 % из воды и заполняющая внутреннюю полость клетки.
В цитоплазме различают две фракции. В одной из них представлены структурные элементы: рибосомы, аэросомы,
карбоксисомы, запасные включения, генетический аппарат, В другой фракции содержится сложная смесь растворимых РНК, ферментных белков, пигментов, минеральных веществ, продуктов и субстратов метаболических реакций. Эта фракция получила название цитозоля.
Благодаря наличию разнообразных органических соединений цитоплазма бактериальных клеток характеризуется повышенной вязкостью. Она в 800-8000 раз больше вязкости воды (приближается к вязкости глицерина). Молодые клетки, находящиеся в лаг-фазе или на начальных этапах логарифмической фазы, имеют более низкую вязкость цитоплазмы; у стареющих - вязкость повышается, напоминая по консистенции гель. Степень вязкости цитоплазмы характеризует не только возраст клетки, но и ее физиологическую активность. Повышение вязкости цитоплазмы у старых культур является одним из факторов, обусловливающих снижение физиологической активности клеток. Цитоплазма является средой, связывающей все внутриклеточные структуры в единую систему.
Рибосомы. В цитоплазме бактериальной клетки постоянно содержатся структуры сферической формы, размером 15-20 нм, молекулярной массой 3106.
Рибосомы состоят на 60-65 % из рибосомальной РНК и на 35- 40 % из белка. Последние богаты основными аминокислотами. При ультрацентрифугировании рибосомы бактерий оседают со скоростью, составляющей около 70 единиц Сведберга (S)7, за что получили название 708-рибосом. Цитоплазматические рибосомы эукариот крупнее и их называют 80S-рибосомами (константа седиментации их равна 80S).
Каждая рибосома состоит из двух субъединиц: 30S и 50S, которые различаются размерами молекул РНК и количеством входящего в их состав белка. Большая субъединица (50S) содержит две молекулы рРНК - 5S и 23S и 35 молекул различных белков. В состав малой субъединицы (30S) входит одна молекула 16 рРНК и 21 молекула разного типа белков. Количество рибосом в клетке непостоянно - от 5000 до 90000. Оно определяется возрастом клетки и условиями культивирования бактерий. Минимальное количество содержится в начале лаг-фазы, а максимальное - в экспоненциальной фазе роста культуры. У кишечной палочки в период активного роста на полноценной питательной среде за 1 с синтезируется 5-6 рибосом. Большая часть их в цитоплазме бактерий находится в свободном состоянии, а остальная -
S = 1 сведберг-единице = 10'13 см (с) ед. поля.
объединена нитями матричной РНК в полисомы. Количество рибосом в полисомах может достигать нескольких десятков. Это свидетельствует о высокой белоксинтезирующей активности клетки, так как рибосомы являются местом белкового синтеза. Их образно называют «фабриками» белка.
Газовые вакуоли (аэросомы). Данные структуры присущи только некоторым водным и почвенным бактериям. Они обнаружены у фототрофных серобактерий, бесцветных нитчатых бактерий, а также у бактерий рода Renobacter. В клетке их содержится до 40-60 (рис. 3.17). Газовые вакуоли окружены тонкой
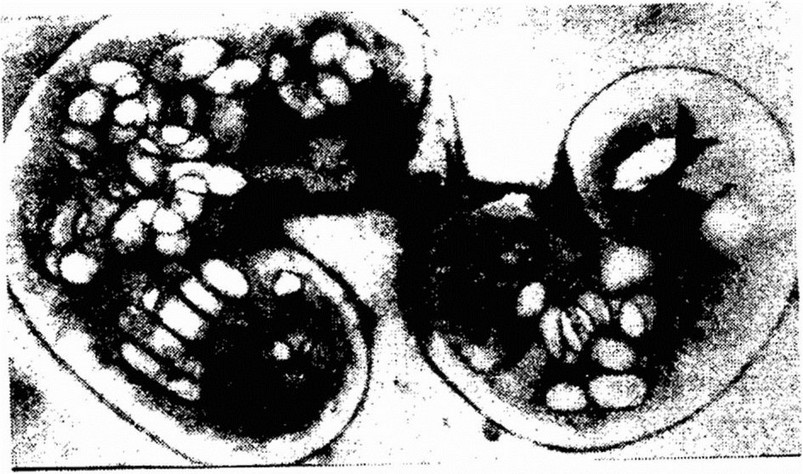
Рис. 3.17. Клетка Renobacter vocuolatum с аэросомами (увеличение х 70 000)
белковой мембраной. Внутри их содержатся газовые пузырьки, число которых непостоянно. Состав и давление газа в пузырьках и аэросомах в целом определяются количеством газов, растворенных в окружающей среде. Аэросомы находятся либо в сжатом состоянии, либо заполнены газом среды. Состояние их регулируется гидростатическим давлением среды. Резкое увеличение давления вызывает сжатие аэросом и клетки при этом утрачивают плавучесть.
Аэросомы регулируют плавучесть клетки, обеспечивая возможность перемещения ее в благоприятные условия аэрации, освещения, содержания питательных веществ. Особенностью является их одноразовое функционирование в состоянии заполненности газом. После сжатия под действием гидростатического давления повторно газом они не заполняются и
постепенно разрушаются. Клетка может воспроизводить их только путем образования заново.
При заполненных газом аэросомах бактерии удерживаются на поверхности воды, при сжатых - погружаются в ее толщу либо оседают на дно водоема. Этот своеобразный способ перемещения выработался в процессе эволюции в основном у бактерий, лишенных жгутиков, а следовательно, и способности к активному передвижению.
Фикобилисомы. Эти внутриклеточные структуры характерны для цианобактерий. Они имеют вид гранул диаметром 28-55 нм, являются местом локализации водорастворимых пигментов - фикобилипротеидов, которые определяю цвет цианобактерий и участвуют в фотосинтезе.
Хлоросомы, или хлоробиум-везикулы - структуры, в которых локализован фотосинтезирующий аппарат зеленых бактерий рода Chlorobium. Они имеют вытянутую форму, длиной 100-150 нм, шириной 50-70 нм, окружены однослойной белковой мембраной. Хлоросомы расположены плотным слоем под цитоплазматической мембраной, но физически отделены от нее. В хлоросомах зеленых бактерий содержатся пигменты фотосинтеза - бактериохлорофиллы, которые поглощают кванты света и энергию передают в реакционные центры фотосинтеза.
Карбоксисомы. В клетках отдельных видов фототрофных (цианобактерии, некоторые пурпурные бактерии) и хемолитотрофных (нитрифицирующие бактерии) прокариот содержатся структуры, имеющие форму многогранника, размером 90-500 нм. В соответствии с выполняемой функцией они получили название карбоксисомы. В них содержится фермент рибулозо- дифосфаткарбоксилаза, который катализирует реакцию связывания углекислоты с рибулозодифосфатом в цикле Кальвина. У автотрофных бактерий они являются местом фиксации двуокиси углерода. Карбоксисомы окружены однослойной белковой мембраной, которая предохраняет фермент от воздействия внутриклеточных протеаз.
Запасные питательные вещества* Кроме описанных структурных элементов, в цитоплазме бактерий содержатся в виде включений гранулы различной формы о размеров. Присутствие их в
клетке непостоянно и связано с составом питательной среды и физиологическим состоянием культуры. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и источником элементов питания. Они образуются обычно в культурах на свежих, богатых питательными веществами средах, когда рост клеток в силу каких-то причин заторможен, или после окончания периода активного роста. Химический состав включений различен и неодинаков у разных видов бактерий. Ими могут быть полисахариды, липиды, кристаллы и гранулы неорганических веществ.
Из полисахаридов следует прежде всего назвать крахмал, гликоген и крахмалоподобное вещество - гранулезу. Наиболее распространенным является гликоген. Он обнаружен у бацилл, сальмонелл, кишечной палочки, сардин и др. У споровых анаэробов рода Clostridium клетки содержат мелкие гранулы гранулезы. Данные включения используются клеткой как источники энергии и углерода.
Липиды накапливаются в цитоплазме бактерий в виде мелких капель и зерен. У многих бактерий липидные включения представлены поли-р-оксимасляной кислотой, на долю которой часто приходится до 50 % сухой биомассы бактерий. Особенно богаты данным соединением бактерии рода Bacillus и фототрофные бактерии. Поли-р-оксимасляная кислота синтезируется в больших количествах при росте микроорганизмов на средах, богатых углеводами. В каждой цепи полилактида на долю остатков р-окси- масляной кислоты приходится до 60 %, в связи с чем для бактерий это соединение является идеальной «кладовой» энергии. У некоторых микроорганизмов накапливаются воска и нейтральные жиры (триглицериды). Так, у микобактерий и актиномицетов воска иногда составляют до 40 % сухой массы, нейтральными жирами богаты клетки дрожжей рода Candida, Rhodotorula, количество их достигает почти 60 %.
Все липидные включения у микроорганизмов служат источником энергии и углерода.
В клетках многих бактерий часто обнаруживаются особые включения, названные зернами вол юти на. По химической природе волютин представляет собой полифосфат. Название волютин
происходит от видового названия серобактерий Spirillum volutans, у которых впервые были описаны эти включения. Волютин обладает свойством метахромазии, т.е. вызывает изменение цвета некоторых красителей. Если бактерии окрасить метиленовым синим или толуидиновым синим, то зерна волютина приобретают пурпурный или красно-фиолетовый цвет. В связи с этим исследователи В. Бабеш и Е. Эрнст, впервые описавшие данные включения, назвали их метахроматичес кими зернами. Зерна волютина имеют сферическую форму, размером до 0,5 мкм. Они образуются в условиях хорошего питания микроорганизмов, особенно на средах, богатых углеводами, а также при наличии в среде глицерина. Обнаруживается волютин в клетках как патогенных, так и сапрофитных бактерий, например, у спирилл, азотобактера, возбудителя дифтерии.
Волютин используется клеткой в основном как источник фосфатных групп и частично энергии.
У бесцветных и пурпурных серобактерий при окислении сульфидов внутри клетки в виде капель откладывается минеральная сера. Накопление серы происходит на средах, богатых сероводородом H2S. При исчерпании сульфидов из среды бактерии используют внутриклеточную серу. Для бесцветных серобактерий она служит источником энергии, для фотосинтезирующих пурпурных серобактерий - донором электронов.
У цианобактерий запасным веществом является цианофицин. Эго полипептид, состоящий из аргинина и аспарагиновой кислоты. Он служит источником азота при недостатке его в среде. Накопление гранул цианофицина происходит в стационарной фазе роста культуры и может составлять до 8 % сухой массы клетки.
Толщина ЦПМ бактерий обычно около 6-8 нм. На ее долю приходится до 15 % сухой массы клетки. Она состоит из липидов (15-45 %), белков (45-60 %) и небольшого количества углеводов (около 10 %). Липиды представлены фосфолипидами - до 30 % сухой массы мембраны. Среди них преобладают фосфатидил- глицерин и дифосфатид ил глицерид (кардиолипин) - обязательный компонент митохондриальных мембран эукариот. В меньшем количестве содержатся фосфатидилинозит и фосфатид ил-
этаноламин. Кроме фосфолипидов в мембране обнаружены различные гликолипиды, небольшие количества каротиноидов и хинонов. В составе липидов, производных глицерина, выявлены нетипичные для мембран жирные кислоты - насыщенные или мононенасыщенные с 16 - 18 углеродными атомами, а также кислоты, не встречающиеся в мембранах эукариот - циклопропановые и разветвленные жирные кислоты с 15-17 углеродными атомами. Набор жирных кислот, как и состоящих из них липидов мембран, является видоспецифичным для прокариот.
Мембранные липиды представлены небольшими полярными молекулами, несущими гидрофильные (головки) и гидрофобные (хвосты) группы. В водной среде они спонтанно образуют замкнутый бимолекулярный слой - бислой. Этот слой служит существенным барьером для ионов и полярных соединений. Организованные в бимолекулярный слой липиды составляют структурную основу мембраны, поддерживают механическую стабильность и придают ей гидрофобность.
Белки составляют больше половины сухой массы мембраны. Их насчитывается более 20 различных типов. Исходя из различий в прочности связи с липидами и расположением в мембране, белки подразделяют на интегральные и периферические. Интегральные белки погружены в гидрофобную область мембраны, где образуют многочисленные связи с углеводородными цепями липидов,
создавая липопротеидные комплексы. Периферические белки локализованы на поверхности гидрофильного слоя и часто присоединяются к интегральным белкам (рис. 3.14).
Рис.3.14. Структура цитоплазматической мембраны: 1 - липиды; 2 - гликопротеиды; 3 - периферические белки; 4 - интегральные белки
Мембранные белки по их функциям в составе мембран можно разделить на две группы: структурные и динамические.
Функции структурных белков ограничиваются поддержанием структурной целостности мембраны. Они располагаются на поверхности гидрофильного липидного слоя, выступая в роли молекулярного бандажа.
К динамическим относят белки, которые непосредственно участвуют во всех процессах, происходящих на мембране. Их разделяют на три класса: транспортные, участвующие в транспорте соединений внутрь и наружу клетки; каталитические, выполняющие функции ферментов в реакциях, происходящих на мембране; белки- рецепторы, специфически связывающие определенные соединения (токсины, гормоны) на наружной стороне мембраны.
Углеводы в мембране находятся не в свободном состоянии, а взаимосвязаны с белками и липидами в гликопротеиды. Они, как
правило, локализованы только на наружной поверхности мембраны и выполняют функции рецепторов узнавания факторов внешней среды.
Цитоплазматическая мембрана бактерий, как и все другие биологические мембраны, является асимметричной жидкокристаллической структурой. Асимметрия обусловлена различиями в химическом строении молекул белка и их расположении в липидном бислое мембраны. Одни белки расположены на поверхности бислоя, другие - погружены в его толщу, третьи проходят насквозь от внутренней до внешней поверхности бислоя. Строго определенная ориентация мембранных белков в свою очередь обусловлена тем, что они синтезируются и включаются в мембрану асимметрично. Наружная и внутренняя поверхности мембраны различаются также по ферментативной активности. В зависимости от условий (например, температуры) ЦПМ может находиться в различных фазовых состояниях: разжиженном или кристаллическом. При переходе одной жидкокристаллической фазы в другую изменяется подвижность компонентов мембраны и плотность ее упаковки, что, в свою очередь, приводит к нарушению ее функциональной активности.
Структурная организация и функции цитоплазматической мембраны. Для объяснения природы и механизма многочисленных функций ЦПМ наиболее подходящей является жидкостно-мозаичная модель организации биологических мембран, предложенная Р. Сингером и А. Николсоном в 1972 г. Согласно данной модели, мембраны представляют собой двумерные растворы определенным образом ориентированных глобулярных белков и липидов. Липиды образуют бислой, в котором гидрофильные «головки» молекул обращены наружу, а гидрофобные «хвосты» погружены в толщу мембраны, обладая при этом достаточной гибкостью. Мембранные липиды и многие белки свободно перемещаются в бислое, но только в латеральном направлении (латеральная диффузия). В поперечном направлении, т. е. от одной поверхности мембраны к противоположной, белки перемещаться не могут, а липиды перемещаются крайне медленно (1 раз за несколько часов). Причиной отсутствия или низкой активности поперечной диффузии, по-видимому, является асимметричное распределение липидов:
одних липидов больше в наружной части бислоя, других - во внутренней. Следствием этого является неодинаковая электронная плотность (проводимость) бислоя в поперечном направлении.
В жидкокристаллическом или разжиженном состоянии ЦПМ находится только при определенных, так называемых
биологических температурах. При понижении температуры ( ниже точки плавления, Тпл) липиды переходят в кристаллическое состояние, повышается степень вязкости вплоть до затвердевания мембраны. Значение температуры, вызывающей затвердевание мембраны, определяется содержанием ненасыщенных и
разветвленных жирных кислот. Чем больше их в мембране, тем ниже температура перехода липидов из жидкокристаллического состояния в кристаллическое.
Прокариоты обладают способностью регулировать текучесть мембраны путем изменения числа двойных связей и длины цепей молекул жирных кислот. Так, у Е. coli при понижении температуры среды от 42° С до 27° С соотношение насыщенных и ненасыщенных жирных кислот в мембране снижается с 1,6 до 1,0, т. е. содержание ненасыщенных жирных кислот достигает уровня насыщенных. Это предотвращает увеличение вязкости и обеспечивает сохранение клетками физиологической активности при пониженной температуре.
ЦПМ выполняет у прокариот многочисленные жизненно важные функции. В основном они определяются локализованными в ней белками, которые выполняют роль каналов, рецепторов, регенераторов энергии, ферментов, транспортные функции и другие. ЦПМ является основным осмотическим барьером, который, благодаря наличию механизмов мембранного транспорта, осуществляет избирательное поступление веществ в клетку и удаление из нее продуктов метаболизма. Избирательная проницаемость ЦПМ обусловлена локализованными в ней субстратспецифическими пермеазами, осуществляющими активный перенос через мембрану различных органических и минеральных веществ. В ЦПМ содержатся ферменты биосинтеза мембранных липидов и макромолекул, входящих в состав клеточной стенки, наружной мембраны, капсулы. ЦПМ является местом локализации окислительно-восстановительных ферментов, осуществляющих
транспорт электронов, окислительное и фотосинтетическое фосфорилирование, генерирование электрохимической энергии
трансмембранного потенциала (А// +) и химической (АТФ). ЦПМ
н
выполняет важные функции в биосинтезе и транслокации секретируемых белков грамотрицательными бактериями. Биосинтез данных белков осуществляется на рибосомах, прикрепленных к ЦПМ. У грамотрицательных бактерий на ЦПМ имеются специальные рецепторные белки, «узнающие» сигналы из большой рибо- сомной субчастицы о прикреплении рибосомы и начале синтеза белка. Мембранные рецепторные белки взаимодействуют с большой субъединицей рибосомы, образуется рибосомомембранный комплекс, на котором осуществляется синтезсекретируемых белков. Таким путем, например, Е. coli синтезирует щелочную фосфотазу, Вас. subtilis - а-амилазу. ЦПМ обеспечивает также перенос данных белков в периплазматическое пространство. Велика роль ЦПМ в регуляции клеточного деления, репликации хромосомы и плазмид и последующей сегрегации этих генетических элементов между вновь образующимися дочерними клетками.
Все прокариоты наряду с цитоплазматической мембраной содержат ее производные - внутриклеточные мембраны, которые выполняют специализированные функции. Цитоплазматическая мембрана способна к образованию всевозможных инвагинаций (впячиваний). Эти инвагинации составляют внутриклеточные мембраны, которые имеют различную протяженность, упаковку и локализацию в цитоплазме. Они могут быть собраны в сложные клубки - пластинчатые, сотовидные или трубчатые образования. Менее сложные мембраны имеют вид простых петель или канальцев различной протяженности. Независимо от сложности организации внутриклеточных мембран, все они являются производными цитоплазматической мембраны. Величина активной поверхности их превышает таковую цитоплазматической мембраны. Это дает основание судить о большой функциональной активности данных структур в клетках.
Особенно богатый внутриклеточный мембранный аппарат обнаружен у азотфиксирующих и фотосинтезирующих бактерий, бруцелл, нитрифицирующих бактерий. У фотосинтезирующих бактерий (Rhodospirillum rubrum) мембраны имеют вид замкнутых пузырьков - везикул. Их образование начинается с впячивания цитоплазматической мембраны, которое затем образует трубочку. На трубочке появляются перетяжки, разделяющие ее на ряд пузырьков. Эти пузырьки называют хроматофорами. В них содержатся поглощающие свет пигменты - бактериохлорофиллы и каротиноиды, ферменты транспорта электронов - убихиноны и цитохромы, компоненты системы фосфорилирования. У некоторых фотосинтезирующих прокариот, в частности у пурпурных серобактерий и цианобактерий, фотосинтезирующий аппарат представлен стопками мембран, которые имеют уплощенную форму и по аналогии с гранами хлоропластов зеленых растений называются тилакоидами (рис. 3.15).
В них концентрируются пигменты фотосинтеза, ферменты электрон- транспортной цепи и системы фосфорилирования. Особенностью тила- коидов цианобактерий является отсутствие связи с цитоплазматической мембраной. Это единственная группа прокариот, имеющая дифференцированную мембранную систему.
У нитрифицирующих бактерий внутриклеточный мембранный аппарат имеет вид пластинок, или ламелл, состоящих из плоских пузырьков (рис. 3.16).
Из внутриклеточных мембран наиболее сложную структуру имеют мезосомы. Они представляют собой спиралевиднозакрученные, плоские или сферически трубчатые тельца. Формируются мезосомы в период клеточного деления в зоне образования поперечной перегородки. Они принимают участие в репликации хромосомы и распределении геномов между дочерними клетками, в синтезе веществ клеточной стенки. На участие
мезосомы в делении клетки указывает связь ее с ДНК нуклеоида. Хорошо развитые мезосомы обнаруживаются только у грам- положительных бактерий.
Накопленные к настоящему времени сведения говорят о том, что мембранные структуры бактерий достаточно дифференцированы и обеспечивают ход различных метаболических процессов в клетке.
- Цитоплазма и цитоплазматические включения
Цитоплазма - полужидкая коллоидная масса, состоящая на 70-80 % из воды и заполняющая внутреннюю полость клетки.
В цитоплазме различают две фракции. В одной из них представлены структурные элементы: рибосомы, аэросомы,
карбоксисомы, запасные включения, генетический аппарат, В другой фракции содержится сложная смесь растворимых РНК, ферментных белков, пигментов, минеральных веществ, продуктов и субстратов метаболических реакций. Эта фракция получила название цитозоля.
Благодаря наличию разнообразных органических соединений цитоплазма бактериальных клеток характеризуется повышенной вязкостью. Она в 800-8000 раз больше вязкости воды (приближается к вязкости глицерина). Молодые клетки, находящиеся в лаг-фазе или на начальных этапах логарифмической фазы, имеют более низкую вязкость цитоплазмы; у стареющих - вязкость повышается, напоминая по консистенции гель. Степень вязкости цитоплазмы характеризует не только возраст клетки, но и ее физиологическую активность. Повышение вязкости цитоплазмы у старых культур является одним из факторов, обусловливающих снижение физиологической активности клеток. Цитоплазма является средой, связывающей все внутриклеточные структуры в единую систему.
Рибосомы. В цитоплазме бактериальной клетки постоянно содержатся структуры сферической формы, размером 15-20 нм, молекулярной массой 3106.
Рибосомы состоят на 60-65 % из рибосомальной РНК и на 35- 40 % из белка. Последние богаты основными аминокислотами. При ультрацентрифугировании рибосомы бактерий оседают со скоростью, составляющей около 70 единиц Сведберга (S)7, за что получили название 708-рибосом. Цитоплазматические рибосомы эукариот крупнее и их называют 80S-рибосомами (константа седиментации их равна 80S).
Каждая рибосома состоит из двух субъединиц: 30S и 50S, которые различаются размерами молекул РНК и количеством входящего в их состав белка. Большая субъединица (50S) содержит две молекулы рРНК - 5S и 23S и 35 молекул различных белков. В состав малой субъединицы (30S) входит одна молекула 16 рРНК и 21 молекула разного типа белков. Количество рибосом в клетке непостоянно - от 5000 до 90000. Оно определяется возрастом клетки и условиями культивирования бактерий. Минимальное количество содержится в начале лаг-фазы, а максимальное - в экспоненциальной фазе роста культуры. У кишечной палочки в период активного роста на полноценной питательной среде за 1 с синтезируется 5-6 рибосом. Большая часть их в цитоплазме бактерий находится в свободном состоянии, а остальная -
S = 1 сведберг-единице = 10'13 см (с) ед. поля.
объединена нитями матричной РНК в полисомы. Количество рибосом в полисомах может достигать нескольких десятков. Это свидетельствует о высокой белоксинтезирующей активности клетки, так как рибосомы являются местом белкового синтеза. Их образно называют «фабриками» белка.
Газовые вакуоли (аэросомы). Данные структуры присущи только некоторым водным и почвенным бактериям. Они обнаружены у фототрофных серобактерий, бесцветных нитчатых бактерий, а также у бактерий рода Renobacter. В клетке их содержится до 40-60 (рис. 3.17). Газовые вакуоли окружены тонкой
Рис. 3.17. Клетка Renobacter vocuolatum с аэросомами (увеличение х 70 000)
белковой мембраной. Внутри их содержатся газовые пузырьки, число которых непостоянно. Состав и давление газа в пузырьках и аэросомах в целом определяются количеством газов, растворенных в окружающей среде. Аэросомы находятся либо в сжатом состоянии, либо заполнены газом среды. Состояние их регулируется гидростатическим давлением среды. Резкое увеличение давления вызывает сжатие аэросом и клетки при этом утрачивают плавучесть.
Аэросомы регулируют плавучесть клетки, обеспечивая возможность перемещения ее в благоприятные условия аэрации, освещения, содержания питательных веществ. Особенностью является их одноразовое функционирование в состоянии заполненности газом. После сжатия под действием гидростатического давления повторно газом они не заполняются и
постепенно разрушаются. Клетка может воспроизводить их только путем образования заново.
При заполненных газом аэросомах бактерии удерживаются на поверхности воды, при сжатых - погружаются в ее толщу либо оседают на дно водоема. Этот своеобразный способ перемещения выработался в процессе эволюции в основном у бактерий, лишенных жгутиков, а следовательно, и способности к активному передвижению.
Фикобилисомы. Эти внутриклеточные структуры характерны для цианобактерий. Они имеют вид гранул диаметром 28-55 нм, являются местом локализации водорастворимых пигментов - фикобилипротеидов, которые определяю цвет цианобактерий и участвуют в фотосинтезе.
Хлоросомы, или хлоробиум-везикулы - структуры, в которых локализован фотосинтезирующий аппарат зеленых бактерий рода Chlorobium. Они имеют вытянутую форму, длиной 100-150 нм, шириной 50-70 нм, окружены однослойной белковой мембраной. Хлоросомы расположены плотным слоем под цитоплазматической мембраной, но физически отделены от нее. В хлоросомах зеленых бактерий содержатся пигменты фотосинтеза - бактериохлорофиллы, которые поглощают кванты света и энергию передают в реакционные центры фотосинтеза.
Карбоксисомы. В клетках отдельных видов фототрофных (цианобактерии, некоторые пурпурные бактерии) и хемолитотрофных (нитрифицирующие бактерии) прокариот содержатся структуры, имеющие форму многогранника, размером 90-500 нм. В соответствии с выполняемой функцией они получили название карбоксисомы. В них содержится фермент рибулозо- дифосфаткарбоксилаза, который катализирует реакцию связывания углекислоты с рибулозодифосфатом в цикле Кальвина. У автотрофных бактерий они являются местом фиксации двуокиси углерода. Карбоксисомы окружены однослойной белковой мембраной, которая предохраняет фермент от воздействия внутриклеточных протеаз.
Запасные питательные вещества* Кроме описанных структурных элементов, в цитоплазме бактерий содержатся в виде включений гранулы различной формы о размеров. Присутствие их в
клетке непостоянно и связано с составом питательной среды и физиологическим состоянием культуры. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и источником элементов питания. Они образуются обычно в культурах на свежих, богатых питательными веществами средах, когда рост клеток в силу каких-то причин заторможен, или после окончания периода активного роста. Химический состав включений различен и неодинаков у разных видов бактерий. Ими могут быть полисахариды, липиды, кристаллы и гранулы неорганических веществ.
Из полисахаридов следует прежде всего назвать крахмал, гликоген и крахмалоподобное вещество - гранулезу. Наиболее распространенным является гликоген. Он обнаружен у бацилл, сальмонелл, кишечной палочки, сардин и др. У споровых анаэробов рода Clostridium клетки содержат мелкие гранулы гранулезы. Данные включения используются клеткой как источники энергии и углерода.
Липиды накапливаются в цитоплазме бактерий в виде мелких капель и зерен. У многих бактерий липидные включения представлены поли-р-оксимасляной кислотой, на долю которой часто приходится до 50 % сухой биомассы бактерий. Особенно богаты данным соединением бактерии рода Bacillus и фототрофные бактерии. Поли-р-оксимасляная кислота синтезируется в больших количествах при росте микроорганизмов на средах, богатых углеводами. В каждой цепи полилактида на долю остатков р-окси- масляной кислоты приходится до 60 %, в связи с чем для бактерий это соединение является идеальной «кладовой» энергии. У некоторых микроорганизмов накапливаются воска и нейтральные жиры (триглицериды). Так, у микобактерий и актиномицетов воска иногда составляют до 40 % сухой массы, нейтральными жирами богаты клетки дрожжей рода Candida, Rhodotorula, количество их достигает почти 60 %.
Все липидные включения у микроорганизмов служат источником энергии и углерода.
В клетках многих бактерий часто обнаруживаются особые включения, названные зернами вол юти на. По химической природе волютин представляет собой полифосфат. Название волютин
происходит от видового названия серобактерий Spirillum volutans, у которых впервые были описаны эти включения. Волютин обладает свойством метахромазии, т.е. вызывает изменение цвета некоторых красителей. Если бактерии окрасить метиленовым синим или толуидиновым синим, то зерна волютина приобретают пурпурный или красно-фиолетовый цвет. В связи с этим исследователи В. Бабеш и Е. Эрнст, впервые описавшие данные включения, назвали их метахроматичес кими зернами. Зерна волютина имеют сферическую форму, размером до 0,5 мкм. Они образуются в условиях хорошего питания микроорганизмов, особенно на средах, богатых углеводами, а также при наличии в среде глицерина. Обнаруживается волютин в клетках как патогенных, так и сапрофитных бактерий, например, у спирилл, азотобактера, возбудителя дифтерии.
Волютин используется клеткой в основном как источник фосфатных групп и частично энергии.
У бесцветных и пурпурных серобактерий при окислении сульфидов внутри клетки в виде капель откладывается минеральная сера. Накопление серы происходит на средах, богатых сероводородом H2S. При исчерпании сульфидов из среды бактерии используют внутриклеточную серу. Для бесцветных серобактерий она служит источником энергии, для фотосинтезирующих пурпурных серобактерий - донором электронов.
У цианобактерий запасным веществом является цианофицин. Эго полипептид, состоящий из аргинина и аспарагиновой кислоты. Он служит источником азота при недостатке его в среде. Накопление гранул цианофицина происходит в стационарной фазе роста культуры и может составлять до 8 % сухой массы клетки.