
Споры у бактерий образуются внутри клетки и носят название эндоспор. Образование эндоспор присуще палочковидным бактериям родов Bacillus и Clostridium и является для них характерным и постоянным признаком. Среди бактерий других морфологических групп спорообразование - редкое явление. Всего известно 15 родов бактерий, отдельные представители которых образуют эндоспоры, например, роды Sporosarcina, Sporospirillum, Desulfotomaculum.
Эндоспоры - это особый тип покоящихся клеток, которые в отличие от цист и акинет образуются внутри вегетативной клетки - спорангия. Споры могут иметь сферическую или овальную форму и занимать в клетке центральное или терминальное положение.
В отличие от вегетативных клеток они обладают высокой устойчивостью к неблагоприятным факторам внешней среды: высокой температуре, высушиванию (годами могут сохраняться в высушенном состоянии), действию повышенных концентраций солей, а некоторые выдерживают даже длительное кипячение. В литературе описаны случаи прорастания спор бактерий, выделенных из гробниц египетских пирамид. Высокая устойчивость спор к неблагоприятным воздействиям имеет большое биологическое значение для спорообразующих бактерий. Эта особенность, а также то, что в клетке, как правило, образуется только одна спора, свидетельствует о том, что процесс спорообразования не является средством размножения бактерий, а служит приспособлением к сохранению вида.
Хотя спорообразование - естественный физиологический процесс, в то же время для спорообразующих бактерий он вовсе не обязательный этап жизненного цикла. При благоприятных условиях питания эти бактерии длительное время могут развиваться без образования спор. Основным фактором, вызывающим спорообразование, является концентрация в среде органических и азотсодержащих веществ. Доказательством этого может быть усиление спорообразования при помещении вегетативных клеток в дистиллированную воду. Через 10-12 ч споры образуют около 90 % клеток. Однако, если в течение первых 5 ч после помещения клеток
в воду добавить глюкозу, то спорообразование прекратится. Благоприятное влияние на образование спор оказывает повышенное содержание кислорода, присутствие в воде катионов марганца и калия. Оптимумы температуры и pH для спорообразования те же, что и для развития бактерий.
Спорообразование - сложный процесс, в котором можно выделить несколько этапов. На первом, подготовительном этапе, изменяется направленность метаболических процессов клетки: происходит активный синтез спорового белка, который в отличие от белка вегетативной клетки содержит больше цистеина и аминокислот с гидрофобными группами; синтезируется уникальное соединение - дипиколиновая кислота, которая присуща только споре. В этот же период происходят морфологические изменения нуклеоида - ядерное вещество вытягивается по длине клетки в виде тяжа. При наличии в клетке двух нуклеоидов, что часто бывает в период завершения экспоненциальной фазы роста, они объединяются и также образуют тяжевидную структуру. Затем часть (примерно 1/3) этой структуры отделяется и переходит к одному из полюсов клетки - в спорогенную зону. Это может быть одна или несколько копий хромосомы материнской клетки. Здесь же происходит уплотнение цитоплазмы, которая вместе с генетическим материалом отделяется перегородкой от остального клеточного содержимого. Перегородка формируется из цитоплазматической мембраны, которая инвагинирует от периферии к центру, срастается и разделяет таким образом спороплазму и цитоплазму клетки (рис. 3.25). Описанные этапы образования спор - обратимы. Если к спорулирующей культуре на стадии формирования проспоры внести антибиотик хлорамфеникол (ингибитор синтеза белков, в том числе и мембранных), то процесс обрастания спороплазмы прекратится и начнется обычный процесс клеточного деления. Дальнейшие этапы спорогенеза необратимы. Следующий этап спорогенеза - образование проспоры. Он состоит в том, что мембрана вегетативной клетки обрастает отделенный участок спороплазмы с ядерным материалом. В результате формируется структура, расположенная внутри материнской клетки и полностью изолированная от нее двумя элементарными мембранами - наружной и внутренней. Эта структура получила название проспоры. В сформированную

стенок. В состав его входят три повторяющиеся субъединицы: мурамовая, аланиновая и тетрапептидная.
С наличием кортекса связаны морфологии и другие свойства споры. Это установлено на мутантах, потерявших способность к формированию кортекса. Дефектные по развитости кортекса споры имели неправильную форму, не преломляли свет, мало
аккумулировали дипиколиновой кислоты и обладали слабой эффективностью прорастания. Степень развитости кортекса определяет уровень обезвоживания спороплазмы и устойчивость споры к действию неблагоприятных факторов. Он предохраняет также спороплазму от действия литических ферментов, разрушающих клетку.
Вслед за формированием кортекса наступает этап образования споровых покровов - оболочки споры, которая может быть многослойной. У разных видов спорообразующих бактерий оболочки различаются по толщине, строению и количеству слоев. Оболочка синтезируется поверх наружной споровой мембраны. В синтезе принимает участие наружная мембрана споры и протопласт материнской клетки. У некоторых бактерий поверх споровой оболочки формируется еще одна структура - экзоспориум. Он может быть однослойным (Cl. botulinum, тип Е), многослойный (С1. corynoforum). Строение его определяется видом бактерий.
Оболочка споры состоит в основном из белка и небольшого количества липидов и гликолипидов. Белок характеризуется наличием значительного количества дисульфидных связей, которые придают высокую механическую прочность наружным покровам, обеспечивают устойчивость к действию литических ферментов и предохраняют спору от преждевременного прорастания. Мутанты спор, лишенные оболочки, прорастают сразу после освобождения из материнской клетки даже в неблагоприятных для роста условиях.
Экзоспориум состоит из белков и липидов. Полагают, что он защищает спору от внешних воздействий.
После завершения формирования оболочек споры начинается ее созревание. Процесс сопровождается активным поглощением уникального соединения - дипиколиновой кислоты, которая синтезируется спорулирующей клеткой только в период спорообразования. В вегетативной клетке дипиколиновая кислота отсутствует. В зрелой споре она представлена в виде хелата кальция - дипиколинат кальция, составляя 10-15 % массы сухой споры. С этим соединением связывают высокую термостабильность спор и пребывание их в покоящемся состоянии. В процессе созревания спора приобретает характерную для вида форму и размеры, занимает соответствующее положение в клетке. Спустя некоторое время происходит автолиз материнской клетки и спора высвобождается в среду.
Спорообразование регулируется генетическим аппаратом клетки. Выявлено более 100 так называемых spo-генов, контролирующих разные этапы формирования спор. Эти гены расположены группами в разных участках хромосомы. При вегетативном росте клетки они не проявляют себя. Полагают, что они находятся под контролем системы репрессии, которая снимается прекращением роста спорообразующей клетки.
Эндоспоры существенно отличаются от образующих их бактерий по химическому составу. Только спорам присуще наличие дипиколиновой кислоты. В отличие от вегетативной клетки споры характеризуются низким содержанием свободной воды. Это одна из причин слабой или предельно низкой активности метаболических процессов спор, а также устойчивости их к воздействию неблагоприятных физико-химических факторов. Долгое время вообще отрицалась какая-либо физиологическая активность спор в состоянии покоя. Только применение совершеннейших микроспирометров позволило установить наличие слабого дыхания спор.
Споры отличаются от вегетативных клеток высоким содержанием липидов. Так, в клеточных стенках Вас. subtilis общих липидов находится до 0,7 % в оболочках споры - 3,0 %. Оболочкам спор свойственно также относительно большое количество белка. Но зато свободных аминокислот в спорах предельно мало: не обнаружена и p-гидрооксимасляная кислота, характерная для вегетативной клетки большинства споровых бактерий. Спорам присуще также сравнительно высокое содержание кальция и магния и несколько меньшее - фосфора и калия.
Споры, как и вегетативная клетка, имеют полный набор ферментов, однако в покоящейся споре они находятся в недеятельном состоянии. Активность ферментов проявляется при
попадании споры в благоприятную среду и при ее прорастании. Прорастанием называется процесс превращения споры в вегетативную клетку. Необходимым условием для этого является наличие влаги.
В прорастании эндоспор, как и их образовании, различают несколько этапов: активацию, инициацию и вырастание.
Освободившиеся из клетки споры даже в благоприятных условиях сразу не прорастают. Остаются в покоящемся состоянии. Но их можно вывести из состояния покоя, подвергнув воздействию активирующих факторов. Таким фактором является повышенная температура - 65-80° С. Кратковременное (5 мин) прогревание споровой суспензии индуцирует способность спор к прорастанию. Но для начала процесса необходим химический пусковой механизм - инициатор прорастания. Роль его могут выполнять углеводы, аминокислоты (в частности L-аланин), неорганические ионы. Прорастание сопровождается сложными физиологическими и биохимическими изменениями.
На первом этапе прорастания происходит гидратация структур, т. е. поглощение воды и набухание, вместе с тем увеличивается проницаемость оболочек, снижается светопреломляемость их.
В результате этого активируются биохимические процессы: усиливается дыхание и возрастает активность литических ферментов. Истинное прорастание начинается на втором этапе, когда под действием литических ферментов растворяются оболочки спор, вначале внутренняя, затем наружная; освобождаются и удаляются дипиколиновая кислота и частично гликопептид. В этот период теряется 25-30 % сухого веса споры. Разрыв оболочек спор обычно происходит в одной любой точке и там появляется «росток» новой клетки. Далее следует удлинение ростка и формирование полноценной вегетативной клетки. Прорастание споры происходит значительно быстрее, чем ее образование. В среднем этот процесс занимает 4-5 ч. Процесс прорастания спор, так же как и образования, носит необратимый характер.
Спорообразование у некоторых видов бацилл сопровождается появлением несвойственных вегетативной клетке соединений. Так, у Вас. thuringiensis - возбудителя паралича у гусениц чешуекрылых насекомых - рядом со спорой образуется белковый кристаллик
ромбовидной формы, получивший название «параспорального тельца» (рис. 3.26). Этот белок обладает высокой токсичностью для личинок некоторых насекомых. Он освобождается из клетки вместе со спорой.

Рис.3.26. Спора и белковый кристалл Вас. thuringiensis
У многих спорообразующих бактерий ранние этапы споруляции сопровождаются образованием различных пептидных антибиотиков - эдеинов, подавляющих синтез ДНК; бацитрацинов - подавляющих синтез клеточных стенок; грамицидинов и полимикс инов, влияющих на структурные и функциональные свойства мембран. Роль этих соединений в спорообразовании известна. Предполагается, что они могут регулировать различные стадии процесса дифференцировки, как образование проспоры, кортекса, оболочек и т. д.
Экзоспоры. Покоящимися клетками и одновременно репродуктивными структурами у актиномицетов наряду с эндоспорами являются экзоспоры. Они образуются путем фрагментации гиф воздушного мицелия на отдельные участки, каждый из которых превращается в спору. Формирование экзоспор сопровождается утолщением клеточной стенки за счет образования дополнительных наружных покровов. Однако такие структуры, как кортекс и экзоспориум, характерные для эндоспор, у экзоспор отсутствуют. Не обнаружено в экзоспорах и дипиколиновой
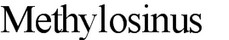

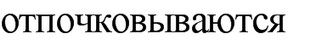
кислоты. В связи с этим, экзоспоры актиномицетов существенно не отличаются от мицелия по устойчивости к повышенной температуре и другим неблагоприятным факторам. Однако в высушенном состоянии они могут более длительное время сохранять жизнеспособность. Обнаружение экзоспор присуще почти всем эуактиномицетам. Кроме актиномицетов экзоспоры образуют некоторые почкующиеся и метанолокисляющие бактерии. У фототрофных бактерий Rhodomicrobium экзоспоры формируются на конце гифы материнской клетки; у метилотрофных бактерий
непосредственно от материнской клетки.