
Жгутики являются нитевидными структурами, расположенными на поверхности клетки (рис. 3.22). Они, подобно капсуле, могут быть удалены без нарушения метаболизма клетки.
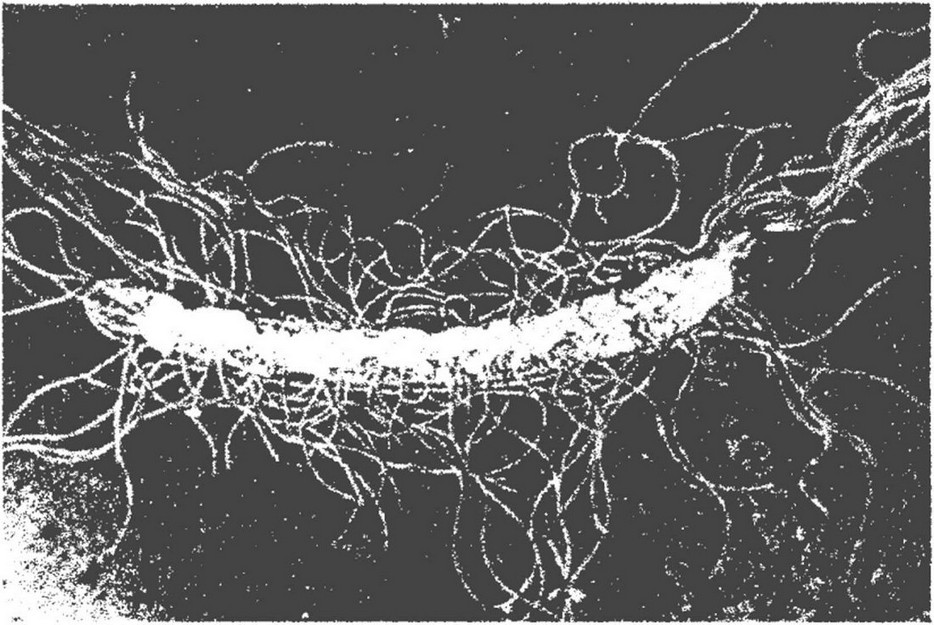
Рис.3.22. Жгутики бактерий
Первые сведения о наличии жгутиков у бактерий сообщил Г. Эренберг в 1838 г., а в 1897 г. В. Мигула дал их морфологическое описание. Количество жгутиков у бактерий непостоянно. Оно изменяется в зависимости от условий культивирования. У спирилл может быть от 5 до 30 жгутиков, у протея - от 50 до 100. Длина жгутиков достигает 20 мкм, что значительно превышает длину бактериальной клетки; диаметр (толщина) жгутиков очень малый - 10-20 нм. По характеру расположения жгутиков бактерии делятся на ряд групп: монотрихи - жгутик на одном полюсе клетки (представители рода Pseudomonas); амфитрихи - по одному жгутику на обоих полюсах (бактерии рода Nitrosomonas); лофотрихи - пучок жгутиков на одном из полюсов (бактерии рода Spirillum); перитрихи - жгутики расположены по всей поверхности клетки (бактерии рода Proteus).
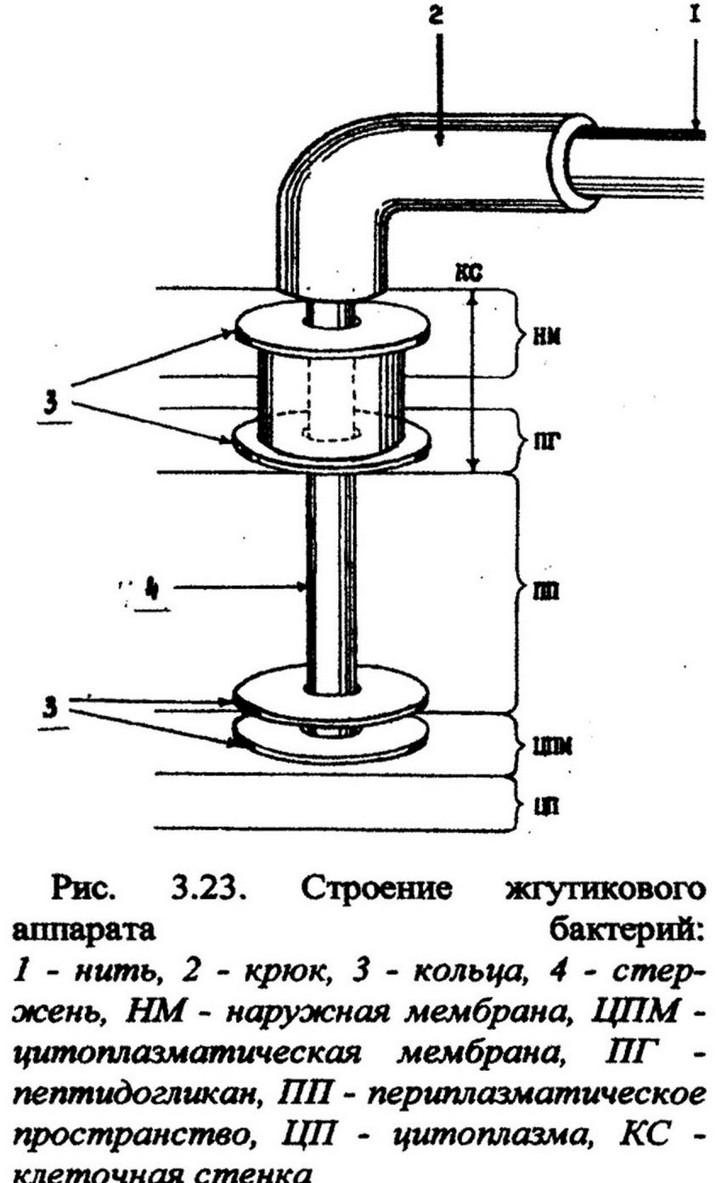
Химический состав жгутиков довольно однообразен. Они состоят из белка флагеллина (от лат. flagellum - жгутик), молекулярная масса которого 25000-60000. В аминокислотном составе флагеллина преобладают глутаминовая и аспарагиновая аминокислоты. Количество ароматических аминокислот незначительное, а триптофан, цистеин и цистин либо содержатся в следовых количествах, либо вовсе отсутствуют. Электронномикроскопические исследования выявили сложную структурную организацию жгутиков. Жгутиковый аппарат состоит из трех частей (рис. 3.23). Основная наружная часть - спиральная жгутиковая нить. У поверхности клеточной стенки она переходит в утолщенное образование изогнутой формы - крюк, который прикрепляет нить к базальному телу, локализованному в ЦПМ и клеточной стенке.
 Нить жгутика состоит из субъединиц белка флагелли- на. У одних бактерий белковые субъединицы собраны в цепи, спирально уложенные по всей длине жгутика с шагом спирали 2-2,5 нм. У других - жгутики имеют фибриллярную структуру - состоят из белковых субфибрилл, которые также имеют спиральную форму укладки, только с большим
Нить жгутика состоит из субъединиц белка флагелли- на. У одних бактерий белковые субъединицы собраны в цепи, спирально уложенные по всей длине жгутика с шагом спирали 2-2,5 нм. У других - жгутики имеют фибриллярную структуру - состоят из белковых субфибрилл, которые также имеют спиральную форму укладки, только с большимшагом спирали. Как в тех, так и других , внутри спирали проходит полый канал, по которому передвигаются молекулы белка из цитоплазмы к дистальному концу (верхушке), с которого идет наращивание жгутика.
Изогнутая часть - крюк - состоит из иного белка, чем нить и полагают, что он служит для обеспечения гибкого соединения нити с базальным телом. В состав базального тела входит сложная система, состоящая из 2 или 4 колец, нанизанных на стержень крючка. У грамположительных бактерий - 2 кольца, обозначенные М и S, у грамотрицательных - 4 - М, S, Р, L. М-кольцо находится в ЦПМ; S - у грамположительных бактерий - в пептидогликановом слое, у грамотрицательных - в периплазматическом пространстве, примыкая к внутренней поверхности пептидогликана. Имеющиеся у грамотрицательных бактерий дополнительные внешние кольца Р и L расположены соответственно в пептидогликановом слое и в наружной мембране. Они обеспечивают более прочное укрепление жгутика в клетках, имеющих тонкий слой пептидогликана и рыхлую клеточную стенку. Внешняя пара колец выполняет функции дополнительной арматуры крепления жгутиков.
Жгутики являются органами движения бактерий. Характер движения определяется особенностью расположения жгутиков. Монотрихи всегда движутся по прямой линии, перитрихи - беспорядочно и с кувырканием. Скорость движения также различна: большинство подвижных форм бактерий за секунду проходит расстояние, близкое размерам их тел. Самая подвижная бактерия - холерный вибрион: при длине тела в 2 мкм он проходит за секунду до 30 мкм.
Перемещение бактерий осуществляется за счет вращательного движения жгутиков. По современным данным жгутики представляют собой спиральные роторы, каждый из которых способен вращаться вокруг собственной оси. Функцию мотора выполняют внутренние кольца М и S. Вращаясь относительно друг друга, они приводят в движение вмонтированный в них стержень крючка, а он, в свою очередь, сообщает вращение связанной с ним жгутиковой нити. Вращение жгутиков передается клетке и она начинает вращаться в противоположном направлении. Спокойное плавательное движение бактерий в одном направлении осуществ-
превращает электрохимическую форму энергии в механическую.
Аналогами бактериальных жгутиков (по строению и химическому составу) являются аксиальные фибриллы (нити) спирохет. Они обвивают протоплазматический цилиндр (тело) клетки, включающий цитоплазму и цитоплазматическую мембрану. Протоплазматический цилиндр окружен наружным чехлом, состоящим из пептидогликана и трехслойной структуры, аналогичной наружной мембране клеточных стенок грамслрица- тельных бактерий. Таким образом аксиальные фибриллы оказываются под чехлом. Число фибрилл разное - от 2 до 100, причем, у каждой клетки два набора фибрилл, прикрепленных субполярно. Свободные концы фибрилл направлены к центру клетки. Спирохеты могут осуществлять движения трех типов: вращение вокруг собственной оси, поступательное винтовое, или штопорообразное, движение и волнообразное изгибание клетки за счет сокращения аксиальных нитей. Движение спирохет, так же как и жгутиковых бактерий, осуществляется за счет электрохимической формы

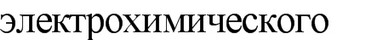
СКОЛЬЗЯ!]
представителям прокариот - миксобактериям, цианобактериям, нитчатым серобактериям также, по-видимому, осуществляется при помощи скрытых внутренних структур - белковых фибрилл. Подтверждением этого является обнаружение у некоторых прокариот со скользящим типом движения тонкого слоя фибрилл, расположенного в клеточной стенке между пептидогликановым слоем и наружной мембраной. У нитчатых цианобактерий фибриллы формируют единую систему, обвивающую в виде спирали весь трихом. Скольжение цианобактерий сопровождается вращением нити. Направление движения зависит от направления хода спирали белковых фибрилл и является таксономическим признаком.
Механизм скользящего движения связан с особенностями строения клеточной стенки, а именно: наличием упорядоченно расположенных белковых субфибрилл. Они аналогичны жгутиковым нитям, только находятся в клеточной стенке, а не на ее поверхности. У скользящих бактерий описаны структуры, напоминающие базальные тела, которые также осуществляют запуск движения фибрилл. В результате их вращательного движения на поверхности клетки появляется так называемая «бегущая волна» -
движение неровной клеточной стенки - и клетка отталкивается от
%
твердого субстрата. Скользящее движение происходит за счет
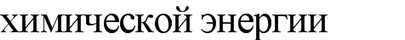
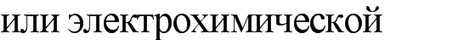 АТФ
АТФ
Полагают, что слизь, обильно выделяемая скользящими прокариотами, в движении их существенной роли не играет. В определенных условиях она может облегчать отталкивание клетки от субстрата.
Подвижные бактерии могут осуществлять направленные передвижения - таксисы, обусловленные различными внешними агентами. В зависимости от характера воздействия (привлечения или отталкивания) таксис бывает положительным или отрицательным. Различают несколько видов таксиса: хемотаксис (реакция клеток на химическое вещество), аэротаксис (на кислород), фототаксис (на свет). Например, фототрофные бактерии благодаря фототаксису скапливаются в освещенном месте; аэробные - в зоне наличия кислорода. Если водную суспензию аэробных бактерий поместить на предметное стекло и покрыть покровным, то через некоторое время масса клеток будет сконцентрирована у края покровного стекла, т. е. там, где более высокое содержание кислорода.