
Первые сведения о ядре бактерий как вполне организованной структуре были получены в 1897 г. благодаря работам М. Мейера. Однако малые размеры бактериальной клетки и высокое содержание РНК, которая окрашивается ядерными красками так же, как и ДНК, затрудняли четкое выявление ядерных структур. Поэтому вопрос относительно наличия ядра у бактерий, его морфологической
структуры и физиологических функций решался в течение многих десятилетий. Не вызывало сомнения наличие у бактерий наследственного аппарата. Это подтверждалось тем, что клетки одного вида бактерий при размножении производят потомство с аналогичными свойствами, т. е. дают культуру исходного вида. Вопрос о ядре у бактерий и его структуре получил окончательное решение только с развитием электронно-микроскопических и генетических исследований. Сейчас установлено, что бактерии имеют структуры, состоящие из ДНК, функционально тождественные ядрам клеток высших организмов. По аналогии они называются бактериальными ядрами или нуклеоидами.
Химическая природа и организация ядерного материала бактерий были установлены австралийским ученым Ж. Кейрнсом в 1963 г. с помощью радиоавтографического метода. Он вносил в питательную среду меченный тритием Н3-тимидин (предшественник тимина) и выращивал на этой среде Е. coli. Затем из клеток бактерий экстрагировалась ДНК, которая помещалась на фотографическую пленку. После соответствующей экспозиции на пленке получался радиоавтограф. Радиоавтография (результат содержания меченого тимина) подтверждала, что исследуемое вещество является дезоксирибонуклеиновой кислотой, ибо ДНК - единственное вещество в клетке, содержащее тимин.
 На радиоавтографе (рис. 3.18) видно, что ДНК Е. coli имеет нитевидную, замкнутую в кольцо структуру, которая реплицируется как единое целое. Кернс зафиксировал последовательные стадии репликации кольцевой ДНК, показав, что обе комплементар-
На радиоавтографе (рис. 3.18) видно, что ДНК Е. coli имеет нитевидную, замкнутую в кольцо структуру, которая реплицируется как единое целое. Кернс зафиксировал последовательные стадии репликации кольцевой ДНК, показав, что обе комплементар-ные нити ДНК удваиваются в точке репликации одновременно.
Длина молекулы ДНК Е. coli составляет 1-1,4 мм. По своим генетическим функциям она тождественна хромосоме.
Таким образом, нуклеоид прокариот представляет собой кольцевую хромосому, которая является гигантской молекулой ДНК с молекулярной массой 1,4-3 х 109 Да. Несмотря на свои относительно крупные размеры бактериальная хромосома - высокоупорядоченная компактная структура. Компактность обеспечивается образованием множества (20-100) суперскрученных петель, которые располагаются в различных областях хромосомы. Бактериальная хромосома взаимодействует в клетке с белками полиамина (спермином и спермидином), которые выполняют функцию, аналогичную гистонам прокариот - нейтрализуют отрицательные заряды ДНК, обусловленные ее химической структурой, а именно, наличием в фосфатных остатках ионизированных гидроксильных групп. Нуклеоид бактерий отличается от ядра эукариотических клеток отсутствием ядерной мембраны, ядрышка и митотического способа деления. Он находится в непосредственном контакте с цитоплазмой клетки.
Репликация ДНК. Одной из функций бактериальной ДНК является репликация (самоудвоение), или воспроизведение себе подобной структуры. Для двухцепочечных кольцевых ДНК характерна двунаправленная репликация. В общих чертах этот процесс можно представить следующим образом. На ДНК (хромосоме) имеются фиксированные точки - локусы, определяющие начало и конец репликации. Эти точки обозначаются буквами «О» ( от origin - начало) и «Т» (termination - окончание) соответственно. Репликация всегда предшествует делению клетки. Хромосома одним или несколькими участками прикрепляется к цитоплазматической мембране. Инициация репликации происходит в точке «Щ» и выражается в появлении репликационных вилок. Цепи ДНК постепенно раскручиваются и каждая из них служит матрицей для образования второй комплементарной цепи. Репликационные вилки продвигаются в противоположных направлениях: одна движется по часовой стрелке, другая - против. По мере их продвижения синтезируются комплементарные цепи ДНК. Обе вилки встречаются в точке окончания репликации («Т»),
которая расположена диаметрально противоположно точке «О» - началу репликации. Репликация заканчивается образованием двух одинаковых молекул ДНК, или двух генетически равнозначных хромосом, несущих одинаковую генетическую информацию, тождественную материнской хромосоме (рис. 3.19). Это
обеспечивается благодаря полуконсервативному механизму
репликации, при котором каждая из образовавшихся молекул ДНК содержит одну родительскую цепь и одну вновь синтезированную.
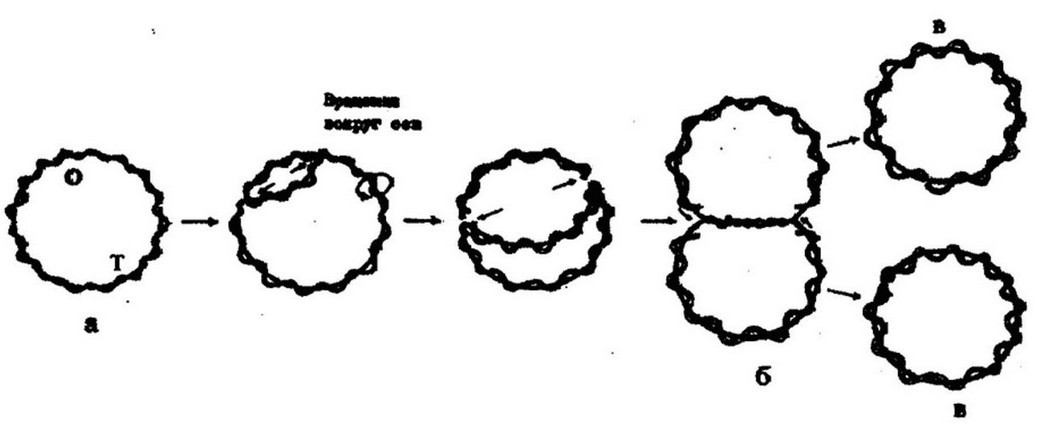
Рис. 3.19. Схема двунаправленной репликации ДНК: а - родительская молекула; б - промежуточные репликативные формы; в - дочерние молекулы; О - точка начала репликации; Т - точка окончания репликации
Репликация ДНК - сложный процесс. В нем принимает участие много разных белков, в том числе и ферментов. Они обеспечивают узнавание точки начала репликации, раскручивание двойной цепи - дуплекса, стабилизацию одиночных цепей, образование затравочной цепи РНК (праймера) для инициации активности ДНК-полимеразы, сборку интактных цепей, узнавание участка терминации, суперскручивание двух новых дуплексов ДНК и образование нативной конформации.
Ведущую роль в репликации ДНК играет ДНК-полимераза. Она связывает между собой нуклеотиды в полинуклеотидную цепь. Причем, связывает только по направлению от 5' к З'-концу. Но так
как ДНК состоит из цепей противоположной полярности (5' gt; У
и 3' gt; 5'), то синтез одной цепи (5' > 3') может происходить
непрерывно в направлении продвигающейся репликационной
вилки, а синтез второй, противоположной цепи (3' gt; 5') должен
идти в обратном направлении. Но ДНК-полимераза неспособна инициировать синтез новой цепи ДНК. Для этого ей необходимо наличие «затравки» - полинуклеотидной цепи со свободным 3-ОН концом. Поэтому синтез ДНК начинается с образования короткого отрезка РНК (10-60 пар оснований), служащего «затравкой», или праймером. Этот процесс обеспечивает фермент ДНК-праймаза, который кеширует часть матричной цепи ДНК. Затем ДНК- полимераза присоединяет свободные нуклеотиды к 3-ОН концу «затравки», образуя короткие отрезки ДНК, так называемые фрагменты Оказаки длиной в 1000-2000 нуклеотидов. По окончании образования всех фрагментов праймер удаляется экзонуклеазой, разрывы между фрагментами застраиваются ДНК-полимеразой в соответствии с матричными участками ДНК. Фрагменты Оказаки сшиваются лигазой, т. е. последовательно соединяются фосфодиэфирными связями. В результате образуются две идентичные двухцепочечные молекулы ДНК (рис. 3.20).
Известны и иные механизмы репликации. Так, удвоение кольцевой ДНК многих вирусов, некоторых фагов и плазмид осуществляется по механизму катящегося кольца. Репликация ДНК у бактерий при конъюгации также происходит аналогичным образом. Это однонаправленный процесс, осуществляющийся следующим образом. В одной из цепей ДНК образуется разрыв и синтез новой цепи начинается с 3'-конца этой разорванной родительской цепи с использованием второй в качестве матрицы. Эго приводит к вытеснению 5'-конца разорванной цепи, которая впоследствии служит матрицей для синтеза новой комплементарной цепи.
Репликация ДНК тесно связана с делением клетки. Расхождение образовавшихся хромосом осуществляется в результате роста клеточной мембраны между точками прикрепления хромосом.
Бактериальный нуклеоид, так же как и ядро клеток растений и животных, является носителем наследственной информации, регулирует направленность белкового синтеза, специфичность белков, и, кроме того, обеспечивает функционирование всех внутриклеточных процессов.
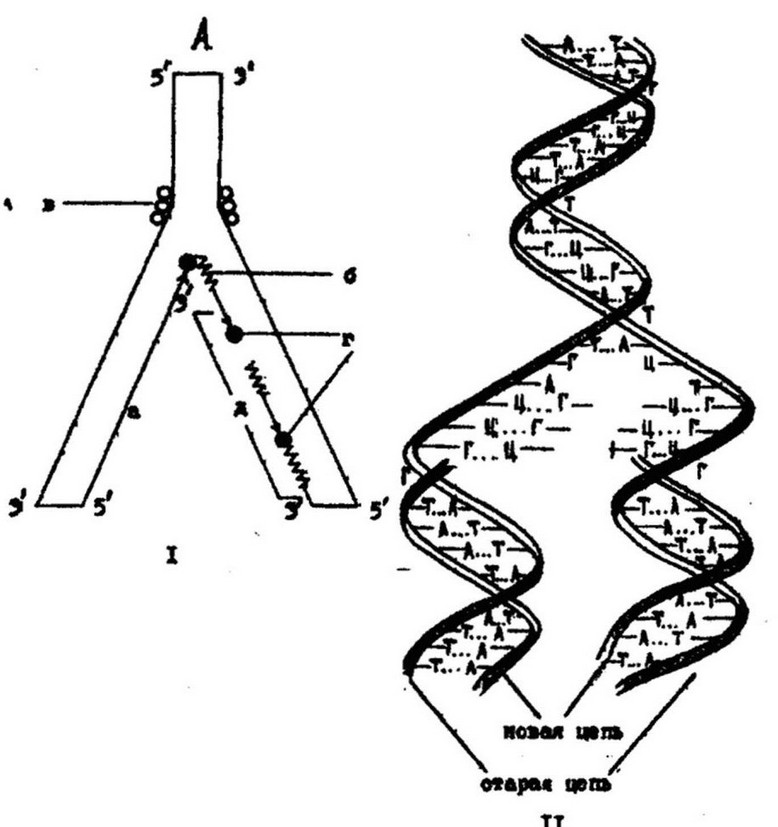
Рис. 3.20. Механизм репликации двуцепочечной ДНК:
I - репликативная вилка; II - полуконсервативный характер репликации; А - старая цепь; а - вновь синтезированная цепь, б - РНК-затравка, в - расплетающие белки, г - ДНК-полимераза, д - фрагменты Оказаки