ЖИРНЫЕ КИСЛОТЫ ЛИПИДОВ ВОДОРОСЛЕЙ
Липиды всех организмов вообще выполняют ряд важнейших жизненных функций, связанных не только с энергообеспечением живой клетки и структурированием ее мембран, но главным образом с синтезом физиологически активных компонентов, необходимых для осуществления фундаментальных процессов в живых системах. Являясь неизменным компонентом мембран клетки и ее органелл, липиды, точнее жирные кислоты, участвуют в обеспечении гомеостаза, очевидно, через включение в биосинтез веществ регуляторного типа действия, в частности простагландинов и их коротко живущих дериватов. Особенно велика биологическая роль полиненасыщенных жирных кислот, яеляющихся субстратом в биосинтезе большого количества эфемерных физиологически активных веществ и кофакторами ряда биологических превращений. И хотя до сих пор полнота функций полиненасыщенных жирных кислот в организмах животных все еще остается невыясненной, она несомненна и весьма существенна для обеспечения нормального функционирования живых систем (Conigalo, 1972). Даже сам факт избирательного накопления полиеновых жирных кислот в жизненно важных органах, а также изменение содержания определенных видов полиненасыщенных жирных кислот в случаях патологических изменений этих органов или при предъявлении к ним чрезмерных нагрузок должен привлечь к этим удивительным эндогенным БАВ пристальное внимание. Удивительным является и то, что из огромного разнообразия существующих в природе жирных кислот, в том числе и высоко непредельных, отдельные органы и ткани биосинтезируют или кумулируют в липидной фракции клеточных мембран строго специфические полиеновые кислоты. Так, например, высшие отделы центральной нервной системы млекопитающих избирательно кумулируют арахидоно- вую и докозагексаеновую кислоты, содержание которых в головном мозгу во много раз превышает концентрацию этих полиенов в других отделах центральной нервной системы и в других органах и тканях. Зато содержание в мозговой ткани других полиненасыщенных жирных кислот, в том числе лино- левой и особенно линоленовой, чрезвычайно незначительно (ли- ноленовая кислота вообще содержится в виде следов). Эти же полиненасыщенные высшие жирные кислоты (арахидоновая, докозагексаеновая) избирательно кумулируются в мышце сердца. Правда, наряду с этими высшими жирными кислотами в мышце сердца определяют заметные количества линолевой кислоты. В тестикулярной ткани, особенно в период функциональной активности, резко возрастает содержание других по- лиеновых высших жирных кислот, например, докозапентаено- вой, докозатетраеновой и т. д., каждая из которых, очевидно, является специфической для определенных видов животных. Полиненасыщенные высшие жирные кислоты служат материалом образования в организмах простагландинов и проста- гландиноподобных веществ. Сами простагландины — это, по существу, ненасыщенные высшие гидроксилированные жирные кислоты с числом углеродных атомов в углеродной цепи, равным 20, которая характеризуется наличием пентацикла в положениях С8-ц.
В литературе считается давно установленным факт участия различных полиненасыщенных жирных кислот в обеспечении ряда физиологических процессов. Это дало повод назвать группу ненасыщенных жирных кислот, включающую, главным образом, линолевую и линоленовую кислоты, витамином Ф. Известно, чо недостаток в пище ненасыщенных жирных кислот (витамин Ф) подобно дефициту в пищевом рационе животных любых других витаминов вызывает достаточно легковоспроизводимые патологические симптомы: понижение плодовитости, склонность к легочным инфекциям, выпадение шерсти, дегенеративные изменения семенных канальцев, поражения кожи, депигментацию, морфологические изменения митохондрий печени, потерю веса и т. д. (Alfin-Slatter, Aftergood, 1968). Для медицины имеет также существенное значение положительное влияние высших полиненасыщенных жирных кислот на функцию печени, миокарда, фибринолитическую активность крови, а также определенное цнтостатическое и особенно гипохолесте- ринемическое действие (Alfin-Slatter, Aftergood, 1968). Исследования показали, что физиологическая активность полиненасыщенных высших жирных кислот возрастает с увеличением ненасыщенное™ соединений. Так, гипохолестеринемический эффект арахндоновой кислоты, имеющей 4 двойные связи в изолированном положении, в среднем в 3,5 раза превосходит таковой линоленовой кислоты, имеющей только 2 двойные связи. Эффект докозагексаеновой кислоты, имеющей 6 изолированных двойных связей, почти в 5 раз превосходит аналогичный эффект линолевой кислоты (Worne, 1959). Особенно благоприятно действует на функцию сосудов и печени, а также уровень холестерина совместное назначение смеси арахндоновой и гексае- новой кислот. Именно это и послужило основой для внедрения в клиническую практику препаратов ненасыщенных высших жирных кислот. Однако спектр этих медицинских препаратов ограничивается, по существу, жирными кислотами, содержащими в основном одну, две и три двойные связи с числом атомов углерода в углеводородной цепи равным 18. Обычным источни-
ком получения этих препаратов являются жиры наземных животных и растений, содержащие значительные количества соединений с 2 и 3 двойными связями. Однако более ценные в биологическом отношении полиеновые высшие жирные кислоты с 4, 5, 6 двойными связями в изолированном положении в наземных животных и растениях встречаются в крайне низких количествах (главным образом арахидоновая кислота). Что касается эйкозапентаеновой (5 двойных связей) и докозаге- ксаеновой (6 двойных связей), то в липидах наземных организмов они определяются в следовых количествах (Ф. М. Ржав- ская, 1976). Зато в липидах морских гидробионтов содержание этих полиеновых соединений достигает у отдельных таксономических видов 30% и более (Ф. М. Ржавская, 1976). Однако изучение биогенеза и транспорта высших полиеновых жирных кислот в морских организмах дает основание считать основными производителями их в Мировом океане различные морские растения, особенно планктонные организмы, являющиеся начальным элементом пищевых цепей в Мировом океане (Г. К. Барашков, 1962). Общее содержание липидов в водорослях зависит от таксономической принадлежности морских растений.
Как показали в 1938 г. исследования Takahashi et al., жирнокислотный состав липидов морских водорослей характеризуется наличием соединений насыщенного и ненасыщенного ряда. Например, в изученных японскими авторами липидах морской водоросли Cystophyllum hakodatense определяются кислоты миристиновая (4,5%), пальмитиновая (18%), гексадеценоевая (16%), гексадекатетраеновая (7%), октадеценоевая (39%), окта- декадиеноевая (3%), октадекатриеноевая (1%), октадекатетрае- новая (7%), докозадиеновая (1%), докозатриеновая (3%) (Takahashi et al., 1938). Значительное количество липидов, включающих как насыщенные, так и ненасыщенные жирные кислоты, содержат планктонные водоросли, отличающиеся также разнообразием положения двойных связей в углеродной цепи. У диатомовой водоросли Sceletonema costatum, характеризующейся относительно высокой концентрацией свободных жирных кислот, последние представлены соединениями, содержащими в основном 2, 3, 4 двойные связи. О жирнокислотном составе липидов этой планктонной водоросли дает представление табл. 7.
Обычными, как свидетельствуют литературные данные, для жирных кислот липидов диатомовых водорослей является 2, 3, 4, 5, 6 двойные связи с числом углеродных атомов в линейной цепи 18, 20 и 22. Общее содержание липидов достигает 30% и более от сухого веса диатомовых водорослей.
Наряду с диатомовыми водорослями синезеленые водоросли (самые распространенные среди микрофитов) также характеризуются значительной биопродукцией липидов (содержание липидов в синезеленых водорослях достигает 10% и более от
Таблица 7

ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ ПЛАНКТОННОЙ
ВОДОРОСЛИ SCELETONEMA COSTATUM
сухого веса водорослей). Главными компонентами липидов синезеленых водорослей аналогично липидам других фитопланктонных организмов являются ненасыщенные жирные кислоты, преимущественно линолевая и линоленовая. О жирнокислотном составе липидных фракций синезеленых водорослей можно составить представление на основании данных, приведенных в таблице 8, из которой видно, что линолевая и линоленовая кислоты вместе составляют около 50% от общего количества жирных кислот исследуемых водорослей.
Что касается других, менее распространенных, классов мик- рофитов, то биопродукция в них липидов отличается такой же выраженностью и направленностью. В качестве примера можно указать на эвгленовые водоросли, липиды которых содержат преобладающее количество ненасыщенных жирных кислот, характеризующихся цис-конфигурацией атомов водорода ’ при двойных связях, что является основной предпосылкой для
Таблица 8
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ НЕКОТОРЫХ ВИДОВ СИНЕЗЕЛЕНЫХ ВОДОРОСЛЕЙ (В МОДИФИКАЦИИ ПО NICHOLS, WOOD, 1968)
Содержание жирных кислот, %
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ EUGLENA GRACILIS И ASTASIA LONGA (В МОДИФИКАЦИИ ПО HULANICKA ЕТ AL., 1964), %
включения их в ряд ферментативных процессов, в частности, процесс биосинтеза ПГ. Исследуя липидные фракции микрофита Euglena gracilis, Hulanicka et al. (1964) определили в них наличие следующих ненасыщенных жирных кислот: 7-гексадецено-
Микроскопические растения, Euglena gracilis и Astasia longa, как видно из результатов опытов, обладают подобно многим фитопланктонным организмам замечательной способностью продуцировать значительные количества высших жирных высоконенасыщенных кислот с 4 и 5 двойными связями в изолированном положении.


Обширный ассортимент жирных кислот идентифицирован также в планктон-
Kalnins 7QK71 у Н°И B0W°PciH Synura Petersenii (Collins, пй17)' X B ЛИПИДП0И Фракции этих микрофитов не обнаружены жирные кислоты, содержащие 4 или даже 3 двой ные связи, тем не менее жир этого гидробионта интересен со Держанием диеновых соединений и жирных кислот с нечетным числом углеродных атомов в цепи. Состав липидной фракции мшфофита Synura petersenii приведен в табл. 10 ФР Несомненно, интенсификация исследования микроскопиче- их морских растений и крупных водорослей с целью иденти-
цеИвтаТскнГ™те^еМеТабОЛИТОВ БАВ’ пРеДставляющих фарма- бпг!Г интеРес, приведет к открытию многих видов _ в а полиненасыщенных жирных кислот. Учиты-
ГГ1 я, Р°СТ0ТУ сбора водорослей и доступность культуры морского фитопланктона, а также несложную технологию изолирования водорослевых липидов, можно надеяться, что в перспективе
производство ненасыщенных жирных кислот из морских водорослей получит значительное развитие. На основе водорослевых липидов можно получать уже в настоящее время как отдельные биологически активные жирные кислоты, так и их эфиры, а также их комплексы, сохраняющие, как известно, присущий по- лиеновым жирным кислотам спектр физиологического действия. Полная экстракция водоросли Euglena gracilis смесью хлороформ—метанол (2:1) с последующим удалением экстрагента, переэтерификацией и газожидкостной хроматографией метиловых эфиров жирных кислот позволяет идентифицировать по крайней мере 35 индивидуальных жирных кислот с числом атомов углерода в цепи более 9, входящих в липидную фракцию жира этого микрофита (Korn, 1964). Наиболее ценную в биологическом отношении фракцию жирных кислот представляют ненасыщенные, особенно содержащие 2 и более двойных связей с числом углеродных атомов в линейной цепи 20 и 22. Именно эти жирные кислоты могут быть использованы для получения различных лечебных препаратов, в том числе и ПГ. Состав и процентное содержание наиболее ценных ненасыщенных жирных кислот липидов Euglena gracilis приведены в табл. 11.
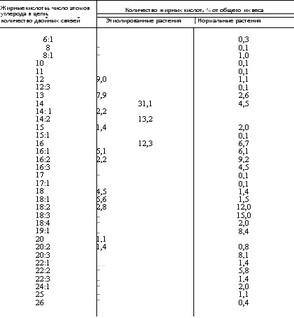
Липиды, несомненно, играют выдающуюся роль в фотосинтезе, причем, как перенос электронов, так и аккумулирование энергии протекает с еще не вполне понятным участием полие- новых кислот, тесно связанных с механизмами клеточного биосинтеза. Именно в этом аспекте следует понимать разительное изменение состава и характера жирных кислот, идентифицируемых в Euglena gracilis при наличии и отсутствии солнечного света (Rosenberg, 1963). Так, при солнечном освещении эти микрофиты накапливают в липидной фракции поли- ненасыщенные жирные кислоты преимущественно с 3 двойными связями и 16, 18, 20 атомами углерода в цепи. В этот период отмечается также усиленное образование еще более непредельных кислот, характеризующихся крайней метастабильностью. Этиолированные растения, наоборот, накапливают в липидной фракции главным образом насыщенные жирные кислоты с числом атомов углерода в цепи менее 17. При этом резко возрастает количество неомыляемых фракций в липидах микрофитов, с 25% при нормальной освещенности до 50% в случае развития в отсутствие солнечного света (Rosenberg, 1963). О влиянии на жирнокислотный состав липидов Euglena gracilis солнечного света можно составить представление по данным определения состава жирных кислот микрофитов, приведенным в табл. 12.
Как известно, существуют значительные видовые различия в жирнокислотном составе липидов различных морских водорослей и это убедительно иллюстрируется данными настоящего раздела монографии, различия, которые обусловлены целым рядом факторов (эволюционно-генетические, условия внешней
среды, возраст водорослей и т. д.). Интересно существующее различие между составом жирных кислот морских и пресноводных водорослей, которое имеет исключительно важное теоретическое и практическое значение. Это различие касается, по существу, наличия или отсутствия в составе жирных кислот соединений, содержащих полиненаеыщенные жирные кислоты с 4, 5, 6 двойными связями в изолированном положении и числом углеродных атомов 20 и 22. Именно полиненаеыщенные жирные кислоты, содержащие 20, 22 углеродных атома в цепи, 4 и более двойных связей (включая Ci84), составляют наиболее ценный в биологическом отношении компонент липидной
фракции водорослей. Эти полиеновые кислоты отсутствуют обычно в липидах пресноводных водорослей, которые в этом отношении уподобляются любым растениям, вегетирующим на суше. В то же время высшие жирные кислоты с 4 и более двойными связями характерны для биосинтетической продукции морских водорослей, являющихся их источниками для практически всех обитателей Мирового океана. Очевидно, механизм биосинтеза полиеновых кислот был частично или полностью утрачен растительными организмами при приспособлении к существованию в пресной воде (и на поверхности суши), в условиях которых возникли иные биогенетические механизмы. Так или иначе для биопродукции пресноводных водорослей полиеновые кислоты, и особенно с 5 и 6 двойными связями, менее характерны, чем для морских водорослей. Учитывая наличие цепей питания, следует ожидать и изменения в жирнокислотном составе пресноводных животных — обитателей рек и озер. И действительно, липиды пресноводных рыб, а также беспозвоночных и млекопитающих (например, животных озера Байкал) содержат значительно меньше полиеновых соединений (за исключением арахидоновой кислоты), чем липиды морских животных (Ф. М. Ржавская, 1976). В качестве иллюстрации приводим результаты сравнительного изучения жнрнокислотного состава липидных фракций ряда морских и пресноводных водорослей (Pohl et al., 1968). В работе авторов были использованы липиды следующих водорослей: Gracilaria confervoides,
Laurencia obtusa, Dictyopteris polydioides, Sebdenia monardiana, Styptocaulon scoparium, Taonia atomaria, Undaria pinnatifida, Ulva fasciata, Valonia urticularis, Halimeda tuna, Platymonas. tetrathele, Codium elongatum, Porhyridium cruentum (морские водоросли, включая планктонные) и Chlorella variegata, Chlorel- la pyrenoidesa, Ankistrodesmus braunii, Scenedesmus obliquus (пресноводные водоросли). Данные исследования состава жирных кислот вышеперечисленных водорослей приводятся в табл. 13.
Анализируя результаты обширного экспериментального исследования морских и пресноводных водорослей, приведенные в табл. 13 и 14, можно со всей определенностью сделать следующие выводы: при прочих равных условиях жирнокислотный состав водорослей определяется их видовой принадлежностью и исторически наличествующей в процессе эволюции средой обитания. При этом морские водоросли в процессе биосинтеза накапливают высшие полиеновые кислоты с числом углеродных атомов больше 18; пресноводные же водоросли заканчивают синтез непредельных жирных кислот на более ранней ступени, накапливая преимущественно жирные кислоты с числом углеродных атомов до 18 и количеством двойных связей до 3. Пресноводные водоросли практически (в данной работе ученых Pohl et al., 1968) не содержат высших полиеновых кислот.
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ МОРСКИХ ВОДОРОСЛЕЙ (ПО POHL ЕТ AL., 1968), %
Эикозатетраеновая (арахидоновая) кислота — основной прекурсор ПГ.
** Эйкозапентаеновая кислота — прекурсор ПГ.
PC — Porphyridium cruentum GC — Gracilaria confervoides LO — Laurencia obtusa SM — Sebdenia monardiana DP — Dictyopteris polypodioides SS — Styptocaulon scoparium ТА — Taonia atomaria UP — Undaria pinnatifida
Это в значительной степени предопределяет направления возможного использования липидов водорослей в медицинских и народнохозяйственных целях. Если липиды морских водорослей представляют собой богатейший источник высоконенасыщенных жирных кислот, содержащих 4 и 5 изолированных двойных связей, то, наоборот, липиды пресноводных водорослей содержат обычные для царства растений суши жирные кислоты, из которых около половины приходится на широко распространенные в растительных маслах (подсолнечное, льняное, кукурузное, хлопковое и т. д.) олеиновую, линолевую и линоле- новую кислоты. Отсюда практически выгодно использовать липиды пресноводных водорослей, в общем случае, для целей
СОДЕРЖАНИЕ ЖИРНЫХ КИСЛОТ В ЛИПИДАХ МОРСКИХ И ПРЕСНОВОДНЫХ ВОДОРОСЛЕЙ (ПО POHL ЕТ AL., 1968), %
* Эйкозатетраеновая кислота — (арахидоновая) — основной прекурсор ПГ.
** Эйкозапентаеновая кислота — прекурсор ПГ.
РТ — Platymonas tetrathela СЕ — Codium elongatum НТ — Halimeda tuna НЕ — Ulva fasciata VU — Valonia utricularis CV — Chlorella variegata CP — Chlorella pyrenoidosa AB — Ankistrodesmus braunii SO — Scenedesmus obliquus
питания, иными словами, вполне рационально использовать пресноводные водоросли на корм скоту, как источник ценных соединений, в том числе легко усваиваемых липидов. В то же время липиды морских водорослей рациональнее применять для изолирования высших полиеновых кислот, особенно арахидоно- вой и эйкозапентаеновой, как основу для создания высокоэффективных и дорогих медицинских препаратов и препаратов для нужд животноводства и звероводства. Во всех случаях морские водоросли содержат в процентном отношении неизмеримо большее количество высших полиеновых кислот, чем любые другие растительные и животные организмы, обитающие на суше. Это делает водорослевые липиды исключительно перспективным сырьем в народном хозяйстве, особенно принимая
 гигантские запасы непрерывно возобновляющейся биомассы морских водорослей. В частности, планктонные водоросли, ответственные за так называемое «цветение» морской воды, «красные приливы», характеризующиеся возможностью безудержного роста на огромных пространствах протяженностью в десятки и даже сотни километров, биосинтезируют липиды, богатые биологически активными высшими полиненасыщенными жирными кислотами. Так, например, фитопланктонный организм, вызывающий периодическое «цветение» морской воды у побережья Северной Калифорнии, Gonyaulax polyedra накапливает в значительных количествах в липидной фракции непредельные жирные кислоты, содержащие 4, 5 и 6 двойных связей в изолированном положении в углеродной цепи, состоящей из 18—22 атомов углерода. Общее количество липидов в Gonyaulax polyedra составляет 30% и более (Patton et al., 1967). Для извлечения липидов микрофиты подсушивают и извлекают несколько раз этанолом и, наконец, смесью (1:1) этанола и эфира. Жирнокислотный состав липидов фракции Gonyaulax polyedra представлен в табл. 15.
гигантские запасы непрерывно возобновляющейся биомассы морских водорослей. В частности, планктонные водоросли, ответственные за так называемое «цветение» морской воды, «красные приливы», характеризующиеся возможностью безудержного роста на огромных пространствах протяженностью в десятки и даже сотни километров, биосинтезируют липиды, богатые биологически активными высшими полиненасыщенными жирными кислотами. Так, например, фитопланктонный организм, вызывающий периодическое «цветение» морской воды у побережья Северной Калифорнии, Gonyaulax polyedra накапливает в значительных количествах в липидной фракции непредельные жирные кислоты, содержащие 4, 5 и 6 двойных связей в изолированном положении в углеродной цепи, состоящей из 18—22 атомов углерода. Общее количество липидов в Gonyaulax polyedra составляет 30% и более (Patton et al., 1967). Для извлечения липидов микрофиты подсушивают и извлекают несколько раз этанолом и, наконец, смесью (1:1) этанола и эфира. Жирнокислотный состав липидов фракции Gonyaulax polyedra представлен в табл. 15.
Обычным путем использования полиненасыщенных жирных кислот морских водорослей является создание на их основе различных препаратов для применения в медицине, животноводстве, звероводстве, птицеводстве. С этой целью после экстракции липидов водорослей их освобождают от растворителей,
обезвоживают и по возможности быстро переэтерифицируют, переводя жирные кислоты в более стабильные эфиры (обычно метиловые или этиловые). Концентраты эфиров жирных кислот обычно используют для получения узких фракций эфиров, например содержащих только С20;4 и Сго:5, или эфиров индивидуальных жирных кислот. Этого можно достичь различными способами, из которых наиболее простым в техническом отношении является метод вакуумной дистилляции. Уже самим эфирам высших полиненасыщенных жирных кислот, особенно арахидоновой, эйкозапентаеновой, докозагексаеновой свойственно выраженное лечебное действие. В отличие от полиеновых кислот их эфиры обладают значительной стойкостью и сохраняют свои свойства при длительном хранении даже в условиях комнатной температуры. Используя эфиры полиеновых кислот как матрицы, возможно вводить в молекулы полиненасыщенных жирных кислот другие активные субстанции, которым было бы желательно сообщить большую лиофильность с целью повышения пенетрирующей способности в отношении цитоплазматических мембран. Обычно в этих целях используют витамины (тиамин, рибофлавин, пантотеновая кислота и др., а также ферменты и химиотерапевтические средства). Таким образом удается создать аддитивно действующий комплекс более активный, нежели простая смесь используемых ингредиентов. Весьма заманчивым представляется применение самих индивидуальных полиеновых кислот, участвующих, как известно, в синтезе многих жизненно важных соединений. Наконец, индивидуальные высшие жирные кислоты с 4, 5, 6 двойными связями могут быть использованы для промышленного получения ПГ и ПГ-подобных веществ. В этом случае рациональнее использовать цис-форму эйкозатетраеновой кислоты (арахидоно- вая кислота), в обычных условиях легко, с помощью специфической ПГ-синтетазы, превращающейся в равные количества важнейших nTF2 и П1Т2а. Также легко вступают в реакции циклизации цис-формы эйкозапентаеновой и докозагексаеновой кислот. Таким образом, и липиды морских водорослей, содержащие значительные количества полиненасыщенных высших жирных кислот, могут найти широкое медицинское и народнохозяйственное применение, что позволит повысить эффективность утилизации биологических ресурсов Мирового океана в интересах всего человечества.
В литературе считается давно установленным факт участия различных полиненасыщенных жирных кислот в обеспечении ряда физиологических процессов. Это дало повод назвать группу ненасыщенных жирных кислот, включающую, главным образом, линолевую и линоленовую кислоты, витамином Ф. Известно, чо недостаток в пище ненасыщенных жирных кислот (витамин Ф) подобно дефициту в пищевом рационе животных любых других витаминов вызывает достаточно легковоспроизводимые патологические симптомы: понижение плодовитости, склонность к легочным инфекциям, выпадение шерсти, дегенеративные изменения семенных канальцев, поражения кожи, депигментацию, морфологические изменения митохондрий печени, потерю веса и т. д. (Alfin-Slatter, Aftergood, 1968). Для медицины имеет также существенное значение положительное влияние высших полиненасыщенных жирных кислот на функцию печени, миокарда, фибринолитическую активность крови, а также определенное цнтостатическое и особенно гипохолесте- ринемическое действие (Alfin-Slatter, Aftergood, 1968). Исследования показали, что физиологическая активность полиненасыщенных высших жирных кислот возрастает с увеличением ненасыщенное™ соединений. Так, гипохолестеринемический эффект арахндоновой кислоты, имеющей 4 двойные связи в изолированном положении, в среднем в 3,5 раза превосходит таковой линоленовой кислоты, имеющей только 2 двойные связи. Эффект докозагексаеновой кислоты, имеющей 6 изолированных двойных связей, почти в 5 раз превосходит аналогичный эффект линолевой кислоты (Worne, 1959). Особенно благоприятно действует на функцию сосудов и печени, а также уровень холестерина совместное назначение смеси арахндоновой и гексае- новой кислот. Именно это и послужило основой для внедрения в клиническую практику препаратов ненасыщенных высших жирных кислот. Однако спектр этих медицинских препаратов ограничивается, по существу, жирными кислотами, содержащими в основном одну, две и три двойные связи с числом атомов углерода в углеводородной цепи равным 18. Обычным источни-
ком получения этих препаратов являются жиры наземных животных и растений, содержащие значительные количества соединений с 2 и 3 двойными связями. Однако более ценные в биологическом отношении полиеновые высшие жирные кислоты с 4, 5, 6 двойными связями в изолированном положении в наземных животных и растениях встречаются в крайне низких количествах (главным образом арахидоновая кислота). Что касается эйкозапентаеновой (5 двойных связей) и докозаге- ксаеновой (6 двойных связей), то в липидах наземных организмов они определяются в следовых количествах (Ф. М. Ржав- ская, 1976). Зато в липидах морских гидробионтов содержание этих полиеновых соединений достигает у отдельных таксономических видов 30% и более (Ф. М. Ржавская, 1976). Однако изучение биогенеза и транспорта высших полиеновых жирных кислот в морских организмах дает основание считать основными производителями их в Мировом океане различные морские растения, особенно планктонные организмы, являющиеся начальным элементом пищевых цепей в Мировом океане (Г. К. Барашков, 1962). Общее содержание липидов в водорослях зависит от таксономической принадлежности морских растений.
Как показали в 1938 г. исследования Takahashi et al., жирнокислотный состав липидов морских водорослей характеризуется наличием соединений насыщенного и ненасыщенного ряда. Например, в изученных японскими авторами липидах морской водоросли Cystophyllum hakodatense определяются кислоты миристиновая (4,5%), пальмитиновая (18%), гексадеценоевая (16%), гексадекатетраеновая (7%), октадеценоевая (39%), окта- декадиеноевая (3%), октадекатриеноевая (1%), октадекатетрае- новая (7%), докозадиеновая (1%), докозатриеновая (3%) (Takahashi et al., 1938). Значительное количество липидов, включающих как насыщенные, так и ненасыщенные жирные кислоты, содержат планктонные водоросли, отличающиеся также разнообразием положения двойных связей в углеродной цепи. У диатомовой водоросли Sceletonema costatum, характеризующейся относительно высокой концентрацией свободных жирных кислот, последние представлены соединениями, содержащими в основном 2, 3, 4 двойные связи. О жирнокислотном составе липидов этой планктонной водоросли дает представление табл. 7.
Обычными, как свидетельствуют литературные данные, для жирных кислот липидов диатомовых водорослей является 2, 3, 4, 5, 6 двойные связи с числом углеродных атомов в линейной цепи 18, 20 и 22. Общее содержание липидов достигает 30% и более от сухого веса диатомовых водорослей.
Наряду с диатомовыми водорослями синезеленые водоросли (самые распространенные среди микрофитов) также характеризуются значительной биопродукцией липидов (содержание липидов в синезеленых водорослях достигает 10% и более от
Таблица 7
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ ПЛАНКТОННОЙ
ВОДОРОСЛИ SCELETONEMA COSTATUM
сухого веса водорослей). Главными компонентами липидов синезеленых водорослей аналогично липидам других фитопланктонных организмов являются ненасыщенные жирные кислоты, преимущественно линолевая и линоленовая. О жирнокислотном составе липидных фракций синезеленых водорослей можно составить представление на основании данных, приведенных в таблице 8, из которой видно, что линолевая и линоленовая кислоты вместе составляют около 50% от общего количества жирных кислот исследуемых водорослей.
Что касается других, менее распространенных, классов мик- рофитов, то биопродукция в них липидов отличается такой же выраженностью и направленностью. В качестве примера можно указать на эвгленовые водоросли, липиды которых содержат преобладающее количество ненасыщенных жирных кислот, характеризующихся цис-конфигурацией атомов водорода ’ при двойных связях, что является основной предпосылкой для
|
Синезеленые водоросли |
||||||
| Жирные
кислоты |
Spirulina
platensis |
Мух оса г- cina
chroococ- coides |
Chlorog-
loea frits ch. |
Anabae- na cylln- drica | Anabaena
flosaquae |
Mastlgo-
cladus laminosus |
Таблица 8
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ НЕКОТОРЫХ ВИДОВ СИНЕЗЕЛЕНЫХ ВОДОРОСЛЕЙ (В МОДИФИКАЦИИ ПО NICHOLS, WOOD, 1968)
Содержание жирных кислот, %
|
16:0 |
43,4 |
38,2 |
42,3 |
46,0 |
39,5 | 38,5 |
|
16:1 |
9,7 |
8,6 |
4,9 |
6,4 |
5,5 42,5 |
|
16:2 |
— |
1,2 |
— |
5,6 |
4,3 — |
|
18:0 |
2,9 |
4,0 |
5,4 |
3,6 |
1,0 — |
|
18:1 |
5,0 |
6,8 |
14,3 |
6,0 |
5,2 16,8 |
|
18:2 |
12,4 |
9,2 |
17,2 |
14,0 |
36,5 2,1 |
|
18:3 |
21,4 |
33,3 |
15,8 |
11,2 |
10,7 - |
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ EUGLENA GRACILIS И ASTASIA LONGA (В МОДИФИКАЦИИ ПО HULANICKA ЕТ AL., 1964), %
|
|
Euglena gracilis |
|
||
|
|
на свету |
|
Astasia |
|
|
Жирные кислоты |
барботаж воздуха с |
в темноте |
Ionga |
|
|
|
5% С02 |
0,5% С02 |
|
|
|
Лауриновая Миристиновая |
0,6 7,2 |
3,5 |
п,б |
5,2 15,2 1,4 16,7 3,8 |
|
Изопентадеканоевая |
|
|
|
|
|
Пальмитиновая 7, 9-Гексадеценовая |
13,9 6,0 |
1Ь,3 4,9 |
19,4 16,0 |
|
|
7, 10-Гексадекадиеновая |
2,8 |
15,8 |
|
|
|
4, 7, 10, 13-Гексадекатетраеновая |
16,0 |
7,1 |
— |
— |
|
Стеариновая |
0,6 |
0,9 |
1,8 |
4,5 |
|
Олеиновая |
10,0 |
10,6 |
7,2 |
7,4 |
|
Линолевая |
3,6 |
15,1 |
2,2 |
1,4 |
|
Линоленовая |
31,5 |
18,8 |
2,2 |
— |
|
11, 14-Эйкозадиеновая |
— |
— |
3,1 |
4,8 |
|
8, 11, 14-Эйкозатриеновая |
|
|
1,1 |
4,4 |
|
Арахидоновая и И, 14, 17-эйкозатриеновая |
3,3 |
3,6 |
10,0 |
17,6 |
|
8, 11, 14, 17-Эйкозатетраеновая |
— |
— |
2,6 |
|
|
5, 8, 11, 14, 17-Эйкозапентаеновая |
2,3 |
2,2 |
9,7 |
4,3 |
включения их в ряд ферментативных процессов, в частности, процесс биосинтеза ПГ. Исследуя липидные фракции микрофита Euglena gracilis, Hulanicka et al. (1964) определили в них наличие следующих ненасыщенных жирных кислот: 7-гексадецено-
- гексадеценовая, 7, 10-гексадекадиеновая, 4,7,10, 13-гексадека- тетраеновая, 11, 14-эйкозадиеновая, 8, 11,14-эйкозатриеновая, арахидоновая, 11,14,17-эйкозатриеновая, 8,11,14,17-эйкозатет- раеновая, 5, 8,11, 14,17-эйкозапентаеновая. Изучение жирнокнс- лотного состава липидов Euglena gracilis показывает, что всякое изменение наиболее важных для мнкрофитов факторов внешней среды, в частности освещения, энергия которого ассимилируется водорослями в процессе фотосинтеза, вызывает изменение направленности биосинтеза жирных кислот и их распределения в клеточных структурах растения. В табл. 9 представлены результаты анализа жирнокислотного состава липидов Euglena gracilis при различной интенсивности освещения культур микрофитов и содержании двуокиси углерода (Hulanicka et al., 1964).
Микроскопические растения, Euglena gracilis и Astasia longa, как видно из результатов опытов, обладают подобно многим фитопланктонным организмам замечательной способностью продуцировать значительные количества высших жирных высоконенасыщенных кислот с 4 и 5 двойными связями в изолированном положении.
Обширный ассортимент жирных кислот идентифицирован также в планктон-
Kalnins 7QK71 у Н°И B0W°PciH Synura Petersenii (Collins, пй17)' X B ЛИПИДП0И Фракции этих микрофитов не обнаружены жирные кислоты, содержащие 4 или даже 3 двой ные связи, тем не менее жир этого гидробионта интересен со Держанием диеновых соединений и жирных кислот с нечетным числом углеродных атомов в цепи. Состав липидной фракции мшфофита Synura petersenii приведен в табл. 10 ФР Несомненно, интенсификация исследования микроскопиче- их морских растений и крупных водорослей с целью иденти-
цеИвтаТскнГ™те^еМеТабОЛИТОВ БАВ’ пРеДставляющих фарма- бпг!Г интеРес, приведет к открытию многих видов _ в а полиненасыщенных жирных кислот. Учиты-
ГГ1 я, Р°СТ0ТУ сбора водорослей и доступность культуры морского фитопланктона, а также несложную технологию изолирования водорослевых липидов, можно надеяться, что в перспективе
производство ненасыщенных жирных кислот из морских водорослей получит значительное развитие. На основе водорослевых липидов можно получать уже в настоящее время как отдельные биологически активные жирные кислоты, так и их эфиры, а также их комплексы, сохраняющие, как известно, присущий по- лиеновым жирным кислотам спектр физиологического действия. Полная экстракция водоросли Euglena gracilis смесью хлороформ—метанол (2:1) с последующим удалением экстрагента, переэтерификацией и газожидкостной хроматографией метиловых эфиров жирных кислот позволяет идентифицировать по крайней мере 35 индивидуальных жирных кислот с числом атомов углерода в цепи более 9, входящих в липидную фракцию жира этого микрофита (Korn, 1964). Наиболее ценную в биологическом отношении фракцию жирных кислот представляют ненасыщенные, особенно содержащие 2 и более двойных связей с числом углеродных атомов в линейной цепи 20 и 22. Именно эти жирные кислоты могут быть использованы для получения различных лечебных препаратов, в том числе и ПГ. Состав и процентное содержание наиболее ценных ненасыщенных жирных кислот липидов Euglena gracilis приведены в табл. 11.
Липиды, несомненно, играют выдающуюся роль в фотосинтезе, причем, как перенос электронов, так и аккумулирование энергии протекает с еще не вполне понятным участием полие- новых кислот, тесно связанных с механизмами клеточного биосинтеза. Именно в этом аспекте следует понимать разительное изменение состава и характера жирных кислот, идентифицируемых в Euglena gracilis при наличии и отсутствии солнечного света (Rosenberg, 1963). Так, при солнечном освещении эти микрофиты накапливают в липидной фракции поли- ненасыщенные жирные кислоты преимущественно с 3 двойными связями и 16, 18, 20 атомами углерода в цепи. В этот период отмечается также усиленное образование еще более непредельных кислот, характеризующихся крайней метастабильностью. Этиолированные растения, наоборот, накапливают в липидной фракции главным образом насыщенные жирные кислоты с числом атомов углерода в цепи менее 17. При этом резко возрастает количество неомыляемых фракций в липидах микрофитов, с 25% при нормальной освещенности до 50% в случае развития в отсутствие солнечного света (Rosenberg, 1963). О влиянии на жирнокислотный состав липидов Euglena gracilis солнечного света можно составить представление по данным определения состава жирных кислот микрофитов, приведенным в табл. 12.
Как известно, существуют значительные видовые различия в жирнокислотном составе липидов различных морских водорослей и это убедительно иллюстрируется данными настоящего раздела монографии, различия, которые обусловлены целым рядом факторов (эволюционно-генетические, условия внешней
среды, возраст водорослей и т. д.). Интересно существующее различие между составом жирных кислот морских и пресноводных водорослей, которое имеет исключительно важное теоретическое и практическое значение. Это различие касается, по существу, наличия или отсутствия в составе жирных кислот соединений, содержащих полиненаеыщенные жирные кислоты с 4, 5, 6 двойными связями в изолированном положении и числом углеродных атомов 20 и 22. Именно полиненаеыщенные жирные кислоты, содержащие 20, 22 углеродных атома в цепи, 4 и более двойных связей (включая Ci84), составляют наиболее ценный в биологическом отношении компонент липидной
фракции водорослей. Эти полиеновые кислоты отсутствуют обычно в липидах пресноводных водорослей, которые в этом отношении уподобляются любым растениям, вегетирующим на суше. В то же время высшие жирные кислоты с 4 и более двойными связями характерны для биосинтетической продукции морских водорослей, являющихся их источниками для практически всех обитателей Мирового океана. Очевидно, механизм биосинтеза полиеновых кислот был частично или полностью утрачен растительными организмами при приспособлении к существованию в пресной воде (и на поверхности суши), в условиях которых возникли иные биогенетические механизмы. Так или иначе для биопродукции пресноводных водорослей полиеновые кислоты, и особенно с 5 и 6 двойными связями, менее характерны, чем для морских водорослей. Учитывая наличие цепей питания, следует ожидать и изменения в жирнокислотном составе пресноводных животных — обитателей рек и озер. И действительно, липиды пресноводных рыб, а также беспозвоночных и млекопитающих (например, животных озера Байкал) содержат значительно меньше полиеновых соединений (за исключением арахидоновой кислоты), чем липиды морских животных (Ф. М. Ржавская, 1976). В качестве иллюстрации приводим результаты сравнительного изучения жнрнокислотного состава липидных фракций ряда морских и пресноводных водорослей (Pohl et al., 1968). В работе авторов были использованы липиды следующих водорослей: Gracilaria confervoides,
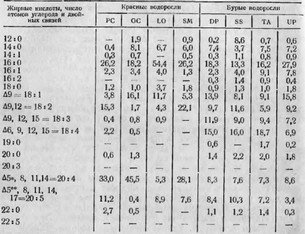
Laurencia obtusa, Dictyopteris polydioides, Sebdenia monardiana, Styptocaulon scoparium, Taonia atomaria, Undaria pinnatifida, Ulva fasciata, Valonia urticularis, Halimeda tuna, Platymonas. tetrathele, Codium elongatum, Porhyridium cruentum (морские водоросли, включая планктонные) и Chlorella variegata, Chlorel- la pyrenoidesa, Ankistrodesmus braunii, Scenedesmus obliquus (пресноводные водоросли). Данные исследования состава жирных кислот вышеперечисленных водорослей приводятся в табл. 13.
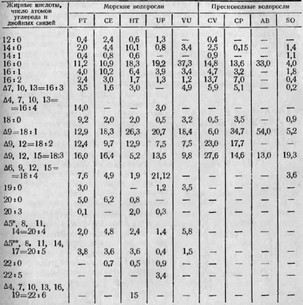
Анализируя результаты обширного экспериментального исследования морских и пресноводных водорослей, приведенные в табл. 13 и 14, можно со всей определенностью сделать следующие выводы: при прочих равных условиях жирнокислотный состав водорослей определяется их видовой принадлежностью и исторически наличествующей в процессе эволюции средой обитания. При этом морские водоросли в процессе биосинтеза накапливают высшие полиеновые кислоты с числом углеродных атомов больше 18; пресноводные же водоросли заканчивают синтез непредельных жирных кислот на более ранней ступени, накапливая преимущественно жирные кислоты с числом углеродных атомов до 18 и количеством двойных связей до 3. Пресноводные водоросли практически (в данной работе ученых Pohl et al., 1968) не содержат высших полиеновых кислот.
ЖИРНОКИСЛОТНЫЙ СОСТАВ ЛИПИДОВ МОРСКИХ ВОДОРОСЛЕЙ (ПО POHL ЕТ AL., 1968), %
Эикозатетраеновая (арахидоновая) кислота — основной прекурсор ПГ.
** Эйкозапентаеновая кислота — прекурсор ПГ.
PC — Porphyridium cruentum GC — Gracilaria confervoides LO — Laurencia obtusa SM — Sebdenia monardiana DP — Dictyopteris polypodioides SS — Styptocaulon scoparium ТА — Taonia atomaria UP — Undaria pinnatifida
Это в значительной степени предопределяет направления возможного использования липидов водорослей в медицинских и народнохозяйственных целях. Если липиды морских водорослей представляют собой богатейший источник высоконенасыщенных жирных кислот, содержащих 4 и 5 изолированных двойных связей, то, наоборот, липиды пресноводных водорослей содержат обычные для царства растений суши жирные кислоты, из которых около половины приходится на широко распространенные в растительных маслах (подсолнечное, льняное, кукурузное, хлопковое и т. д.) олеиновую, линолевую и линоле- новую кислоты. Отсюда практически выгодно использовать липиды пресноводных водорослей, в общем случае, для целей
СОДЕРЖАНИЕ ЖИРНЫХ КИСЛОТ В ЛИПИДАХ МОРСКИХ И ПРЕСНОВОДНЫХ ВОДОРОСЛЕЙ (ПО POHL ЕТ AL., 1968), %
* Эйкозатетраеновая кислота — (арахидоновая) — основной прекурсор ПГ.
** Эйкозапентаеновая кислота — прекурсор ПГ.
РТ — Platymonas tetrathela СЕ — Codium elongatum НТ — Halimeda tuna НЕ — Ulva fasciata VU — Valonia utricularis CV — Chlorella variegata CP — Chlorella pyrenoidosa AB — Ankistrodesmus braunii SO — Scenedesmus obliquus
питания, иными словами, вполне рационально использовать пресноводные водоросли на корм скоту, как источник ценных соединений, в том числе легко усваиваемых липидов. В то же время липиды морских водорослей рациональнее применять для изолирования высших полиеновых кислот, особенно арахидоно- вой и эйкозапентаеновой, как основу для создания высокоэффективных и дорогих медицинских препаратов и препаратов для нужд животноводства и звероводства. Во всех случаях морские водоросли содержат в процентном отношении неизмеримо большее количество высших полиеновых кислот, чем любые другие растительные и животные организмы, обитающие на суше. Это делает водорослевые липиды исключительно перспективным сырьем в народном хозяйстве, особенно принимая
гигантские запасы непрерывно возобновляющейся биомассы морских водорослей. В частности, планктонные водоросли, ответственные за так называемое «цветение» морской воды, «красные приливы», характеризующиеся возможностью безудержного роста на огромных пространствах протяженностью в десятки и даже сотни километров, биосинтезируют липиды, богатые биологически активными высшими полиненасыщенными жирными кислотами. Так, например, фитопланктонный организм, вызывающий периодическое «цветение» морской воды у побережья Северной Калифорнии, Gonyaulax polyedra накапливает в значительных количествах в липидной фракции непредельные жирные кислоты, содержащие 4, 5 и 6 двойных связей в изолированном положении в углеродной цепи, состоящей из 18—22 атомов углерода. Общее количество липидов в Gonyaulax polyedra составляет 30% и более (Patton et al., 1967). Для извлечения липидов микрофиты подсушивают и извлекают несколько раз этанолом и, наконец, смесью (1:1) этанола и эфира. Жирнокислотный состав липидов фракции Gonyaulax polyedra представлен в табл. 15.
Обычным путем использования полиненасыщенных жирных кислот морских водорослей является создание на их основе различных препаратов для применения в медицине, животноводстве, звероводстве, птицеводстве. С этой целью после экстракции липидов водорослей их освобождают от растворителей,
обезвоживают и по возможности быстро переэтерифицируют, переводя жирные кислоты в более стабильные эфиры (обычно метиловые или этиловые). Концентраты эфиров жирных кислот обычно используют для получения узких фракций эфиров, например содержащих только С20;4 и Сго:5, или эфиров индивидуальных жирных кислот. Этого можно достичь различными способами, из которых наиболее простым в техническом отношении является метод вакуумной дистилляции. Уже самим эфирам высших полиненасыщенных жирных кислот, особенно арахидоновой, эйкозапентаеновой, докозагексаеновой свойственно выраженное лечебное действие. В отличие от полиеновых кислот их эфиры обладают значительной стойкостью и сохраняют свои свойства при длительном хранении даже в условиях комнатной температуры. Используя эфиры полиеновых кислот как матрицы, возможно вводить в молекулы полиненасыщенных жирных кислот другие активные субстанции, которым было бы желательно сообщить большую лиофильность с целью повышения пенетрирующей способности в отношении цитоплазматических мембран. Обычно в этих целях используют витамины (тиамин, рибофлавин, пантотеновая кислота и др., а также ферменты и химиотерапевтические средства). Таким образом удается создать аддитивно действующий комплекс более активный, нежели простая смесь используемых ингредиентов. Весьма заманчивым представляется применение самих индивидуальных полиеновых кислот, участвующих, как известно, в синтезе многих жизненно важных соединений. Наконец, индивидуальные высшие жирные кислоты с 4, 5, 6 двойными связями могут быть использованы для промышленного получения ПГ и ПГ-подобных веществ. В этом случае рациональнее использовать цис-форму эйкозатетраеновой кислоты (арахидоно- вая кислота), в обычных условиях легко, с помощью специфической ПГ-синтетазы, превращающейся в равные количества важнейших nTF2 и П1Т2а. Также легко вступают в реакции циклизации цис-формы эйкозапентаеновой и докозагексаеновой кислот. Таким образом, и липиды морских водорослей, содержащие значительные количества полиненасыщенных высших жирных кислот, могут найти широкое медицинское и народнохозяйственное применение, что позволит повысить эффективность утилизации биологических ресурсов Мирового океана в интересах всего человечества.
А так же в разделе « ЖИРНЫЕ КИСЛОТЫ ЛИПИДОВ ВОДОРОСЛЕЙ »
- ВОПРОСЫ КЛАССИФИКАЦИИ
- ПОЛИЕНОВЫЕ КИСЛОТЫ
- ТЕРПЕНОИДЫ
- АЗОТСОДЕРЖАЩИЕ АЛИФАТИЧЕСКИЕ СОЕДИНЕНИЯ
- СОЕДИНЕНИЯ РЯДА БЕНЗОЛА И ИХ ПРОИЗВОДНЫЕ
- ПЕРСПЕКТИВЫ ПРИМЕНЕНИЯ БАВ ГИДРОБИОНТОВ В МЕДИЦИНЕ
- ГАЛОГЕНСОДЕРЖАЩИЕ СОЕДИНЕНИЯ — САМАЯ МНОГОЧИСЛЕННАЯ ГРУППА бав гидробионтов
- МОРСКАЯ ФАРМАЦИЯ — НОВАЯ ОТРАСЛЬ ФАРМАЦЕВТИЧЕСКОЙ НАУКИ
- МОРСКИЕ ВОДОРОСЛИ — НОВЫЙ ИСТОЧНИК БАВ
- СТЕРИНЫ
- ГАЛОГЕНСОДЕРЖАЩИЕ ХИМИЧЕСКИЕ СОЕДИНЕНИЯ
- К ОЦЕНКЕ АНТИБИОТИЧЕСКОЙ АКТИВНОСТИ МОРСКИХ ВОДОРОСЛЕЙ