
При кратком описании реакций клеточного иммунитета, представленном выше, упоминалось, что первичное распознавание чужеродного антигена осуществляется функционально неподготовленными, наивными, Т-клетками, хотя внутритимусная диф- ференцировка уже определила основные свойства этих клеток, что отражается в их делении на субпопуляции.
Созревание наивных Т-клеток в зрелые, функционально активные (армированные) Т-клетки происходит в лимфоидной ткани. Место проникновения патогена (в более широком смысле — антигена) в организм, как правило, удалено от лимфоидных об
разований. Стоком лимфы антиген попадает в ближайшие лимфа- тические узлы. Если антиген оказывается в кровотоке* то он оседает в селезенке. Локализация антигена на слизистых приводит к его миграции в лимфоидную ткань* ассоциированную с этими слизистыми покровами* — в пейеровы бляшки* миндалины,
В периферической лимфоидной ткани имеются три типа специализированных клеток, которые способны усваивать антиген и представлять его в иммуногенной форме на своей поверхности для распознавания Т-клетками, Это — макрофаги, дендритные клетки и В-клетки (табл. 9.1, рис. 9.7). Все они получили общее название антигенпрезентирующнх клеток (АПК). Макрофаги не имеют гистологически определенного места локализации и широко представлены по всей лимфоидной ткани. Дендритные клетки связаны с Т-зоной лимфатических узлов, В-клетки концентрируются в фолликулах. Функция этих типов клеток — представление антигенных пептидов в комплексе с молекулами I или 11 класса* т.е. придание проникшему антигену иммуногенных свойств.
Оказавшийся в лимфоидной ткани антиген провоцирует усиление рециркуляции лимфоцитов. Наивные Т-клетки попадают в лимфатические узлы в так называемую Т-зону (см. главу 6) через высокий эндотелий венул. Генерация зрелых (армированных) эффекторов Т-клеточного иммунного ответа начинается с распознавания антигенного пептида* комплексироваиного с молекулами I или II классов МНС, на поверхности макрофагов и дендритных клеток. Сам факт распознавания комплекса является обязательным, но недостаточным условием для инициации развития наивных Т-клеток в зрелые эффекторы. Необходимо предупреждающее включение кофакторов, которые способствуют взаимодействию рецептора Т-клеток с антигенным комплексом. Именно ан- тигенпрезентирующие клетки обеспечивают такое двойное взаимодействие.
Активация наивных Т-клеток при первичной встрече с антигеном получила название примирование. Это определение введено с тем, чтобы отличить первичное взаимодействие с антигеном от взаимодействия зрелых эффекторов с тем же антигеном, когда собственно и проявляется функциональное предназначение различных субпопуляций Т-клеток. В ряде случаев, в частности при формировании специфической, цитотоксической реакции, пре- зентирующая антиген клетка может выступать и как объект распознавания, и как объект цитолитического действия CD8 Т-клеток после их созревания (см. представленные выше рисунки). Среди наивных Т-лимфоцитов, проникающих в лимфоидную ткань и временно локализующихся в Т-зонах, только один из I05 оказывается способным к специфическому взаимодействию. Остальные постепенно перемещаются в медуллярную зону и через эфферентный лимфатический сосуд покидают орган, чтобы вновь оказаться в кровотоке и быть занесенным в другие лимфоидные образования организма.
9.L2.2. Антигенпрезентирующие клетки в процессе активации наивных Т-клеток
Выше отмечалось (глава 7), что хоминг наивных Т-клеток в лимфатические узлы осуществляется в результате взаимодействия
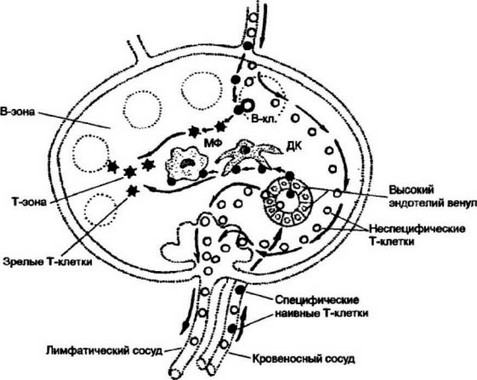
Рис. 9.7. Рециркуляция Т-клеток после проникновения антигена в лимфоидную
ткань.
Антиген с током лимфы от места проникновения в организм заносится в лимфатический узел. Антигенпреэентирующие клетки (макрофаги — МФ, дендритные клетки — ДК, В-клетки — В-кл.) захватывают, перерабатывают и представляют антигенные пептиды в комплексе с молекулами I или II классов МНС на своей поверхности. Эти события активируют процесс рециркуляции клеток. Рециркулирующие наивные Т-клетки, среди которых имеются и специфичные к комплексу лимфоциты (черные кружки) попадают в лимфатический узел либо по афферентным лимфатическим сосудам, либо по кровеносным сосудам. В тех случаях, когда антиген поступает в орган с кровотоком, ему приходится преодолевать высокий эндотелий венул. Оказавшись в паренхиме органа, наивные антигенспеш- фи чес кие Т-клетки после распознавания иммуногена на одной из антиген презен- тирующих клеток остаются в органе, с тем чтобы пройти дополнительную диффе- ренцировку до функционально зрелых, армированных CD8- и CD4 Т-клеток. Ан- тигенспецифические клетки, оказавшись невостребованными, покидают орган по эфферентному лимфатическому сосуду и вступают таким образом в новый цикл рециркуляции
L-селектинов Т-клеток с муииноподобными адрессинами сосудов (CD34, GlyCAM-1). При хоминге лимфоцитов в лимфоидную ткань слизистых в процесс включаются L-селектины и MAdCAM-1 эн* дотелия слизистых покровов. Через эндотелий в паренхиму органа Т-клетки проникают за счет другой группы адгезивных молекул — интегрина LFA-1 и членов суперсемейства иммуноглобулинов ICAM-1, -2, -3 (табл. 9.2).
Таблица 9.2
Адгезивные молекулы, участвующие в заселении лимфоидной ткани наивными Т-клеткнми и генерации зрелых Т-клеток
|
Процесс |
Эндотелий венул |
Антигснпре- зентируюшие клетки |
Т-клетки |
|
Заселение лимфоидной ткани |
CD34, GlyCAM-l, MAdCAM-1, lCAMs |
|
L-селектин LFA-1 |
|
Взаимодействие наивных Т-клеток с антиген пре- |
|
ICAMs |
LFA-1 |
|
зентирующими клетками |
|
LFA-3 (CD58) |
CD2 |
Проникновение наивных Т-лимфоцитов в кортикальную зону приводит к их столкновению с антигенпрезентирующими клетками. На начальном этапе процесса, как это ни странно, отсутствует специфический компонент взаимодействия, т.е. взаимодействие Т- клеточного рецептора с иммуногеном на поверхности антигенп- резентирующих клеток. В межклеточный контакт вступают LFA-1 и CD2 Т-клеток, а также ICAM-1, -2, -3 и LFA-3 антигенпрезен- тирующих клеток. Дифференциальная роль каждого из этих адге- зинов не установлена. Возможно, что их одновременная экспрессия на клеточной поверхности обеспечивает определенный “запас прочности”. Известно, что отсутствие синтеза LFA-1 у людей не нарушает функционирования Т-системы. Выпадение звена взаимодействия пары LFA-1:1CAM компенсируется более активным включением пары CD2:LFA-3.
Преходящее взаимодействие наивных Т-клеток с антигенпрезентирующими клетками осуществляется низкоаффинными LFA-1. Отсутствие жесткой формы контактных отношений между клетками имеет конкретный биологический смысл. Низкая аффинность адгезина позволяет Т-клетке приходить в контакт со многими антигенпрезентирующими клетками до тех пор, пока не произойдет специфическое узнавание иммуногена соответствующим антигенраспознающим рецептором. Как только наивная Т-клетка находит “свой” иммуноген, ее перемещение в корковом слое приостанавливается. От Т-клеточного рецептора идет сигнал на LFA-1, меняющий конформацию адгезина, что приводит к усилению его аффинности по отношению к молекулам ICAM (рис.
- . Механизм изменения аффинности LFA-1 не известен. Вероятно, в этом процессе принимают участие ионы Mg+2, но не Са+2. Возникшие изменения стабилизируют контактные отношения между антигенспецифическими наивными Т-клетками и антигенпре- зентирующими клетками. В результате такая Т-клетка становится подготовленной к пролиферации и дифференцировке в зрелые ан- тигенспецифические Т-клетки.
Подавляющее большинство наивных Т-клеток не выдерживает отбора на специфичность и покидает лимфатический узел через эфферентный лимфатический сосуд, с тем чтобы вновь вступить в процесс рециркуляции в поисках соответствующих по специфичности антигенов (см. рис. 9.7).
Связывание антигенспецифического рецептора с комплексом пептид:молекулы I или II класса МНС и включение в комплексообразован ие корецепторов CD8 или CD4 обеспечивает лишь одно из условий развития наивных Т-клеток — формирование первого сигнала к пролиферации и дифференцировке этих клеток. Чтобы специфически подготовленная клетка начала, наконец, процесс дальнейшего развития, необходим второй сигнал от клеточной поверхности к геному (рис. 9.8). Костимулятором в данном случае выступает молекула В7, экспрессирующаяся на мембране анти- ген-презентирующей клетки. В7 — это гомодимер, относящийся к суперсемейству иммуноглобулинов. Его костимулирующая активность была продемонстрирована в опытах по переносу гена для этого белка в фибробласты, которые сами по себе не относятся к антигенпрезентирующим клеткам. Внесение в культуру Т-клеток, получивших специфический сигнал об антигене, фибробластов с трансфецированным геном для В7, обеспечивало полноценное развитие специфического ответа. Рецептором дтя В7 на поверхности наивной Т-клетки является белок CD28, также относящийся к суперсемейству иммуноглобулинов. Взаимодействие CD28:B7 обеспечивает формирование второго сигнала. Антитела к В7, препятствующие его взаимодействию с CD28, отменяют эффект ко- стимуляции.
Помимо CD28 активированные наивные Т-клетки синтезируют и экспрессируют еще один белок с костимулирующей активностью — CTLA-4. Он обладает большей аффинностью по отношению к В7. Между CD28 и CTLA-4 высока гомология по последовательности аминокислотных остатков, а кодирующие их гены близко сцеплены в хромосоме. Ясно, что совместное костимули-
рующее действие этих молекул оказывает больший эффект на развитие Т-клеток.
Тот факт, что одна и та же антигенпрезентирующая клетка выполняет двойную функцию — представление антигена в им-
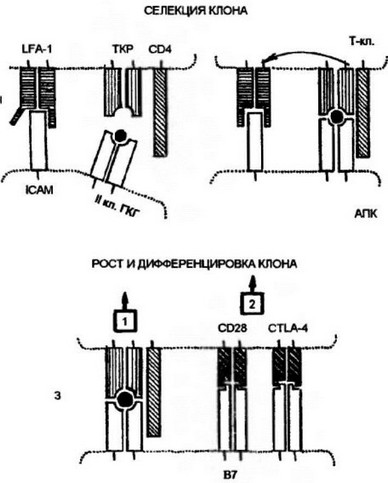
Рис. 9.8. Участие адгезивов и костммуляторов в отборе и дифференцировке наивных Т-клеток.
I. Проникшие в кортикальную зону наивные Т-лимфоциты взаимодействуют неспецифически с антиген представляющими клетками (АПК) посредством адгеэинов: LFA-1 — на Т-клстках и ICAM — на АКП. Взаимодействие LFA-1 — 1САМ на этом этапе характеризуется низкой аффинностью. Если Т-клеточный антигенраспознающий рецептор (ТКР) “не находит” на АКП соответствующего по специфичности комплекса пептид:молекула МНС (пример генерации Т-хслпе- ров), то клетка покидает АПК и вступает в процесс рециркуляции. 2. При соответствии по специфичности ТКР комплексу лептид:молекула МНС повышается аффинность взаимодействия LFA-1 с ICAM, и контактное взаимодействие Т-клетки с АПК становится достаточно прочным. 3. Взаимодействие ТКР с антигенным комплексом формирует при участии корсцептора (на рис. корецептор Т-хелперов CD4) первый сигнал к дифференцировке провзаимолействовавших клеток. Второй сигнал формируется костимулятором В7, представленным на поверхности АПК, и соответствующими лигандами на Т-клетках — CD28 и CTLA-4. Только при совместном действии первого и второго сигналов происходят рост и дифференцировка наивных Т-клеток до зрелых эффекторов
муногенной форме и экспрессию лиганда для костимуляции, т.е. определяет формирование специфического и неспецифического сигналов для дифференцировки наивных Т-клеток в активные эффекторы, имеет большой биологический смысл. Отрицательная селекция в тимусе, очевидно, не является абсолютно безошибочным процессом. Определенные “запрещенные” клоны могут выйти в циркуляцию и стать потенциальной причиной аутоиммунного поражения. Однако, как правило, этой аутоагрессии не наблюдается, поскольку сам факт распознавания антигена, в том числе и собственного, не является единственным условием запуска дифференцировки наивных Т-клеток. Необходимо включение кости- мулятора, которое происходит только на антигенпрезентирующих клетках.