
Участие хеяперных CD4 Т-клеток в продукции антител
Впервые необходимость кооперации Т- и В-лимфоцитов при развитии гуморального иммунного ответа продемонстрировали Клэмен с соавт. в 1966 г. (рис. 9.16). Введение летально облученным мышам клеток только костного мозга (источника В-клеток) или
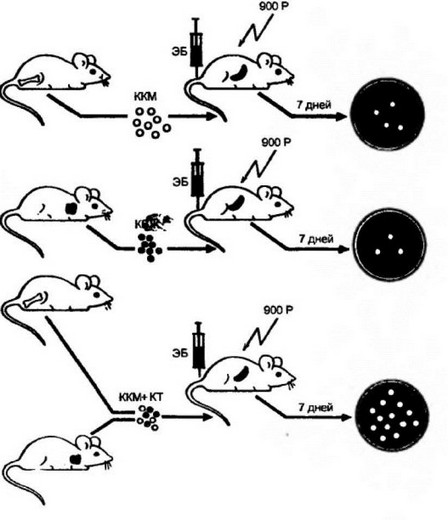
Рис. 9.16. Взаимодействие между клетками тимуса (источника Т-клеток) и клетками костного мозга (источника В-клеток) при индукции гуморального иммунного ответа (Clamp;man et а)., 1966),
Введение облученным мышам клеток только костного мозга (ККМ) или только тимуса (КТ) не обеспечивает развитие иммунного ответа достаточной силы. В то же время введение смеси этих клеток приводит к интенсивной продукции антител к использованному для иммунизации антигену (эритроцитам барана — ЭБ). Причем ответ при совместной инъекции клеток значительно выше, чем сумма ответов при раздельном введении клеток различного происхождения. Таким образом, кооперация двух типов клеток приводит к синергическому эффекту. Ответ оценивали по количеству бляшкообразующих (антителообразуюших) клеток (БОК) в селезенке
только тимуса (источника Т-клеток) не приводило к развитию гуморального иммунного ответа. Продукция антител находилась на крайне низком уровне. В то же время при одновременной инъекции смеси этих клеток наблюдался сильный иммунный ответ. При одновременном введении различных типов клеток он был значительно выше, чем сумма ответов при раздельной инъекции. Иначе говоря, кооперация Т- и В-клеток дает выраженный синергический эффект. Именно синергизм при клеточном взаимо- 240
действии вызвал наибольший интерес исследователей. Эти опыты определили создание целого направления исследований, задача которого — познание механизмов клеточного взаимодействия при развитии иммунного ответа.
Поверхностный иммуноглобулин (slg) играет двойную роль при трансформации В-клеток в антителопродуцирующие плазмо- циты. Во-первых, подобно антигенраспознающему рецептору Т- клеток он передает сигнал о встрече с антигеном непосредственно внутрь клетки. Во-вторых, служит молекулярным фактором захвата этого антигена для переноса его в цитоплазму клетки. Там антиген разрушается до отдельных пептидов, которые в комплексе с молекулами II класса МНС выносятся на поверхность клетки. Иммуногенный комплекс является объектом распознавания ан- тигенспецифическими хелперными CD4 Т-клетками (Тн2), которые обеспечивают второй сигнал для активации В-клеток (см. рис. 9.9; 3).
Тимусзависимые и тимуснезависимые антигены Для большинства антигенов взаимодействие хелперных Т-клеток с В-лимфо- цитами является обязательным условием развития гуморального иммунного ответа. В силу этого подобные антигены (в основном белки, включающие бактериальные токсины) получили название тимусзависимых антигенов (англ, сокр.: “TD antigens”).
Животные или человек с дисфункцией тимуса не способны к полноценной продукции антител. При подобном иммунодефи- цитком состоянии В-клетки не получают второго сигнала от Т- клеток и переходят в состояние анергии (рис. 9.17).
Хотя для индукции антителогенеза к большинству антигенов требуются зрелые Т-клетки, многие компонентны микробов, такие как бактериальные полисахариды, липополисахарады, высокополимерные белки, могут включать В-клетки без дополнительной помощи со стороны хелперных Т-клеток. Эта категория антигенов получила названгие тимуснезависимых антигенов (англ, сокр.: “TI antigens”).
Тимуснезависимые антигены подразделяются на два класса, которые активируют В-клетки разным способом. TI-1-антигены содержат характеристику, которая проявляется в прямой индукции пролиферации В-клеток. При высокой концентрации антигена в процесс пролиферации и дифференцировки вступят все В-клетки, независимо от специфичности их антигенраспознающих рецепторов. Такие антигены получили название поликлональных активаторов, или В-клеточных митогенов. Примером в данном случае может служить липополисахарид бактерий. Снижение концентрации TI-антигена на несколько порядков приводит к стимуляции только антигенспецифического клона В-клеток. Концентрационный эффект связан с наличием у TI-1 антигенов так называемого
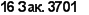 24!
24!
Рис. 9.17. Отлет В-клеток на тнмусзависимый и тимуснсзавксимый антигены.
1. В условиях, когда В-клетки (В) провзаимодейспювали с антигеном (АГ). но не получили помощи со стороны хелперных CD4 Т-клеток (Т), развивается функциональное подавление клетки — анергия. 2. При адекватном включении в ответ хелперных CD4 Т-клеток и формировании в результате двух сигналов для В- клеток (первый — от поверхностного иммуноглобулина (slg), провзаимодейство- вавшего с антигеном (АП, второй — от иммуногенного комплекса на В-клетках) создаются условия для полноценного развития В-клеток в антителопродуиенты. Антигены, которым необходима помощь со стороны Т-клеток, получили название тимуезависимых антигенов. 3. В-клеточный ответ может развиваться без помощи со стороны Т-клеток в тех случаях, когда он формируется на тимуснезависи- мые антигены (TI). Известно два класса таких антигенов. К первому классу (ТЫ) относятся антигены, в чью структуру включат так называемый митогенный участок (МУ), который формирует второй сигнал для В-клеток и тем самым заменяет помощь со стороны Т-клеток. Второй класс — тимуснезависимыс антигены (П-2) — представлен соединениями, в структуре которых имеются повторяющиеся гомологичные эпитопы. Многоточечное соединение этих эпитопов с поверхностными иммуноглобулинами оказывается достаточным для включения В-клстки в процесс пролиферации и дифференцировки до зрелого антителопродуцента митогенного участка, который заменяет сигнал от хелперных Т- клеток. При высоких концентрациях антигена, когда нет конкуренции за специфическое взаимодействие между поверхностным иммуноглобулином и эпитопом TI-1 антигена, наблюдается поликлональный ответ. В условиях, когда концентрация антигена незначительна, преимущество остается за клетками, способными к двойному распознаванию — как митогенного участка, так и антигенного эпитопа. В результате наблюдается клональная экспансия антигенспецифических В-клеток. Для Т1-1 антигена не требуется многократного повторения идентичных эпитопов, так как нали-
чие митогенного участка компенсирует отсутствие многоточечного взаимодействия, что имеет место при активации В-клеток TI - 2-антиге н ам и.
В течение обычного инфекционого процесса концентрация TI-1 -антигенов незначительна. В силу этого будут активироваться только те В-клетки, которые имеют соответствующий антиген- распознающий рецептор, а продуцируемые антитела будут взаимодействовать только с TJ-1 -антигеном. Такой ответ развивается к ряду внеклеточных инфекций, причем он формируется быстрее, чем Т-заьисимый иммунный ответ, так как не требует времени на созревание антигенспецифических хелперных Т-клеток. Скорость ответа сопряжена с несовершенством самого ответа. Он действительно напоминает “скорую помощь” в самом начале развития инфекции. TI-1-антигены, действуя самостоятельно, не способны индуцировать переключение синтеза антител с одного изотипа на другой, не оказывают влияния на повышение аффинности антител, не создают клеток памяти. Все эти процессы требуют хелперных Т-клеток.
Второй класс тимуснезависимых антигенов представлен молекулами, имеющими часто повторяющиеся идентичные эпитопы. К этому классу относятся полисахариды бактериальной стенки, сильно полимеризованные высокомолекулярные белки (например, гемоцианин). Механизм активации В-клеток связан с перекрестным сцеплением между повторяющимися антигенными эпитопами и поверхностным иммуноглобулином. Является ли ответ к TI-i-антигенам полностью независимым от хелперных Т- клеток, не ясно. С одной стороны, гуморальный иммунный ответ регистрируется у дефицитных по Т-клеткам мышей nude. С другой — удаление из культуры in vitro всех Т-клеток приводит к подавлению ответа на TI-2-антиген. Введение незначительного количества Т-клеток иммунодефицитным мышам значительно усиливает иммунный ответ к данным антигенам. Вероятно, наиболее ранний ответ в виде синтеза IgM формируется без помощи Т-клеток. Более поздний гуморальный ответ, характеризующийся доминантным синтезом IgG, требует включения хелперных Т-клеток.
Сцепленное распознавание. В условиях конкретной вирусной или бактериальной инфекции В- и Т-клетки должны распознать тот же самый комплексный антиген, но не обязательно те же самые эпитопы этого антигена. Понимание того факта, что В- и Т- клетки могут реагировать с разными эпитопами и что при этом помощь со стороны хелперных Т-клеток проявляется в полной мере, пришло из опытов по индукции гуморального иммуного ответа у животных, иммунизированных комплексом гаптенгноси- тель (рис. 9.18). Мышей иммунизировали конъюгатом NIP-OA (N1P — гаптен 4-гидрокси-З-йод- 5-нитрофенилуксусная кислота; ОА
243
Таблица 9.4
Сравнительная характеристика различных классов антигенов, инициирующих ^моральный иммунный ответ
— овальбумин кур, использованный в качестве носителя для гап- тена). При переносе клеток селезенки примированных мышей в организм сингенного облученного хозяина развивается анти-NlP- ответ только после введения того же антигена NIP-OA. Продукция антител к гаптену отсутствовала, если в качестве антигена использовали NIP, конъюгированный с другим носителем, например с бычьим сывороточным альбумином (BSA) или сывороточным альбумином человека (HSA). Отсутствовал ответ и в тех случаях, когда перед введением реципиенту клеток селезенки освобождали от Т-клегок, используя анти-Thy-l -сыворотку. Эта первая часть опытов демонстрировала, во-первых, необходимость примированных Т-клеток для развития гуморального иммунного ответа, и во-вторых, распознавание разных эпитопов NIP-OA конъюгата. Если бы Т-клетки распознавали тот же гаптен, то его введение на ином носителе не отразилось бы на силе гуморального ответа. В последующих вариантах этих опытов проводили адаптивный перенос примированных клеток селезенки от мышей иммунизированных как NIP-OA, так и только посторонним носителем — BSA. В последнем случае иммунный ответ развивался и к NIP-OA, и к NIP- 244
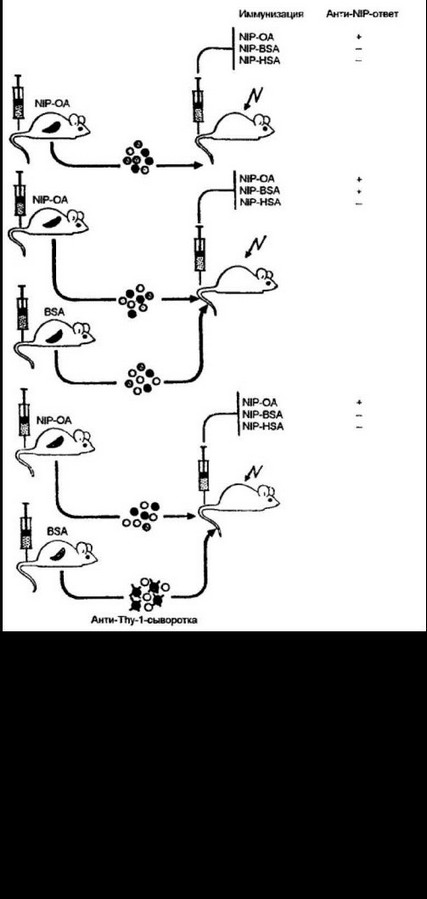
BSA. Удаление Т-клеток из селезенки мышей, иммунизированных BSA, отменяло и продукцию антител к гаптену. Ясно, что имму- ногенность конъюгата связана с возможностью Т-клеток взаимодействовать с носителем. При этом антигенные свойства носителя не имеют значения для специфичности гуморального иммунного ответа.
При реальной бактериальной или вирусной инфекции, как и в эксперименте, Т- и В- клетки распознают разные эпитопы одного и того же комплексного в антигенном отношении патогена. Это явление распознавания В- и Т-клетками разных эпитопов одного и того же антигена получило название сцепленного распознавания. На примере вирусной инфекции цепь событий выглядит следующим образом. Поверхностные белки вирусных частиц, взаимодействуя с предетерминированными антигенраспознающими рецепторами (slg) В-клеток, оказываются поглощенными этими В-клетками. В результате внутриклеточной переработки на клеточной поверхности оказываются пептиды вирусных белков в комплексе с молекулами II класса МНС. Комплекс образуется с пептидами не только внешних вирусных белков, но и внутренних, недоступных для поверхностных иммуноглобулинов В-клеток. Таким образом, В-клетка оказывается подготовленной к встрече со зрелыми хелперными Т-клетками. Т-хелперы проходят свой путь при- мирования, взаимодействуя с тем или иным типом антигенпре- зентирующих клеток. Среди Т-клеток будут те, которые способны реагировать с иммуногенными комплексами, включающими пептиды как внешних, так и внутренних белков. Таким образом помощь В-клеткам усиливается способностью Т-клеток распознавать множество пептидных вирусных фрагментов, но при этом специфичность секретируемых В-ютеткой антител будет только к поверхностному белку вируса, так как первоначальная встреча В- лимфоцитов была именно с поверхностными эпитопами. Образовавшиеся вирусспецифические антитела обладают как нейтрализующей, так и опсонизирующей активностью, т.е. с одной стороны, препятствуют взаимодействию вируса с клеткой-мишенью, а с другой — усиливают контакт с фагоцитирующими клетками через Fc-фрагмент антител.
Мембранные и секреторные активаторы В-клеток. Зрелые хел- перные CD4 Т-клетки, распознавшие комплекс антигенного пептида с молекулами II класса МНС, активируют В-клетки к пролиферации и дифференцировке в зрелые антителопродуценты — плазмоциты. Хелперные Т-клетки после взаимодействия с В-лим- фоцитами начинают экспрессировать мембраносвязанные и секреторные формы молекулярных активаторов (рис. 9.19). Наиболее важными эффекторными молекулами хелперных Т-клеток на ранней стадии активации В-клеток является лиганд CD40, относя- 246
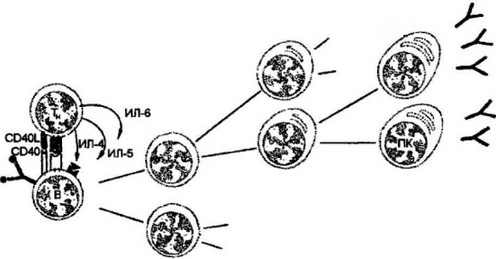
Рис. 9.19. Активация В-клеток к пролиферации и дифференцировке в плазмоци-
ты.
В-клетки (В), экспрессирующие комплекс антигенный пептид:молекула II класса МНС, взаимодействуют с антигснспе пифически ми хелпсрными Т-клетка- ми ГГц). При этом формируются дополнительные сигналы, необходимые В-клетке для выхода в дифференцировку: первый сигнал — от взаимодействия CD40 В- клеток с лигандом CD40L Т-клетсж и второй сигнал — от секретируемого Т- хелперамицитокина интсрлейкин-4 (ИЛ-4). Два дополнительных цитокина — интерлейкин-5 (ИЛ-5) и интерлейкин-6 (ИЛ-6), секрета русмы е Т-клеткам и, вносят свой вклад в процесс дифференцировки В-клеток на заключительных этапах развития
щийся к семейству ФНО-цитокинов. Этот лиганд связывается с поверхностной молекулой В-клеток CD40 — аналогом рецептора к ФНО-ct на макрофагах. Вторым стимулирующим В-клетки фактором является интерлейкин-4 (ИЛ-4), секреция которого хелпер- ными Т-клетками начинается после их контакта с В-лимфоцита- ми. Как и в случае с инфицированными макрофагами, требующими для своей активации двух молекулярных сигналов, синтезируемых Т-клетками воспаления (поверхностного ФНО-а и секрети- руемого цитокина ИНФ-у), В-клетки также активируются двумя стимуляторами — поверхностным СО40-лигандом и секретируе- мым ИЛ-4. Однако если активация инфицированных макрофагов приводит к быстрому эффекторному действию — разрушению внутриклеточных патогенов, то начальный этап активации наивных В- клеток связан с рядом подготовительных процессов, и в первую очередь с их клональной экспансией — накоплением антигенспе- цифического клона, который после завершения пролиферации переходит к дифференцировке в плазматические клетки, секрети- рующие антитела заданной специфичности. Два дополнительных цитокина — ИЛ-5 и ИЛ-6, секретируемые хелпервыми Т-клетками, вносят свой вклад в процесс дифференцировки В-клеток на заключительном этапе процесса В-клсточной активации.
Переключение синтеза изотипов. По мере развития гуморального иммунного ответа синтез антител переключается с одного изотипа на другой при сохранении их исходной специфичности. В самом начале иммунного ответа доминирующим изотипом является IgM. В очень незначительном количестве представлен IgD. Позднее, по мерс развития антителогенеза происходит замена синтеза IgM на синтез IgG, IgA и IgE. Эти изменения в преимущественном синтезе того или иного изотипа не наблюдаются у индивидуумов с дефектом экспрессии СО40-лиганда, принимающего участие в контакном взаимодействии хелперных Т-клеток с В-лимфо- иитами. При подобной форме иммунодефицита наблюдается необычно высокое содержание IgM в сыворотке крови на фоне отсутствия продукции иммуноглобулинов иных изотипов. Активная продукция IgM при недостатке Т-клеточной помощи может быть связана с ответом В-клеток на тимуснезависимые антигены. В то же время эти клинические наблюдения дополнительно демонстрируют, что для сбалансированной продукции различных классов антител необходимо включение в ответ хелперных Т-клеток.
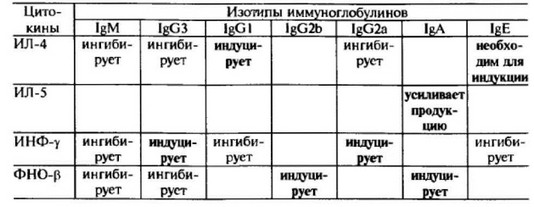
Большинство из того, что известно о регуляции переключения изотипов, пришло из экспериментов, проведенных с мышиными В-клетками, которые стимулировались очищенными цитокинами in vitro. Эти эксперименты показали, что различные цитокины преимущественно индуцируют переключение синтеза “своих” изотипов. Так, ИЛ-4 индуцирует переключение синтеза IgM на IgGI и lg, в то время как ФНО-р — на IgG2b и IgA. Другой цитокин — ИЛ-5, секретируемый хелперными CD4 Т-клетками, усиливает продукцию IgA, когда переключение в геноме для синтеза данного изотипа уже произошло. Хотя CD4 Т-клетки воспаления являются плохими регуляторами синтеза антител, они принимают участие в переключении изотипов посредством секрети- руемого ими ИНФ-у. Этот цитокин оказывает преимущественное влияние на индукцию синтеза IgG3 и IgG2a.
Данные об участии различных цитокинов в продукции антител разных изотипов суммированы в табл. 9.5.
Таким образом, участие хелперных Т-клеток в гуморальном иммунном ответе имеет двойное проявление: во-первых, они инициируют пролиферацию и дифференцировку распознавших антиген В-клеток и, во-вторых, оказывают прямое действие на переключение синтеза антител с одного изотипа на другой.
Повышение аффинности антител в процессе формкрования.им- муиного ответа. По мере развития иммунного ответа происходит не только переключение синтеза антител с одного изотипа на другой, но и повышается аффинность (сродство) этих антител к тому антигенному эпитопу, который вызвал их образование. Константа диссоциации “ранних” антител на два-три порядка выше, чем у
Таблица 9.5
Влияние цитокинов на экспрессию различных изотипов
иммуноглобулинов
“поздних” антител. Кроме того, поданным изоэлектофокусирова- ния известно, что на раннем этапе иммунного ответа гетерогенность антител одной специфичности выше, чем на поздних этапах процесса.
Одна из гипотез, пытающихся объяснить это явление, строится на представлении о гипермутабелыюсти генов, контролирующих вариабельные домены иммуноглобулинов. Известно, что мутационный уровень для всех соматических клеток составляет замену одной пары оснований на 1012 пар при одном клеточном делении. По гипермутационпой теории частота замен представляется величиной, равной одной замене на 103 пар оснований. При этом гипермутабельность не затрагивает гены, контролирующие константные регионы. Первое и основное возражение в отношении данной гипотезы заключается в том, что представление о выборочной повышенной мутабельности каких-то участков ДНК (в данном случае V-генов) кажется умозрительным и не укладывается в современные концепции молекулярной генетики. Непонятно, зачем строить искусственные конструкции, когда уровень исходной генетически детерминированной вариабельности 2,4* 10* разных по специфичности иммуноглобулинов более чем достаточен для взаимодействия с самыми различными антигенными эпитопами; среди них будут и такие, взаимодействие с которыми осуществляется с наибольшей аффинностью.
Вопрос разрешается достаточно просто на основании представлений о существовании сыекции антигенспецифических В- ююнов. На рис. 9.20 показан один из возможных механизмов повышения аффинности антител в процессе развития иммунного ответа зрелыми В-клетками. В реакцию распознавания антигенного эпитопа вступают клетки нескольких клонов, имеющих анти-
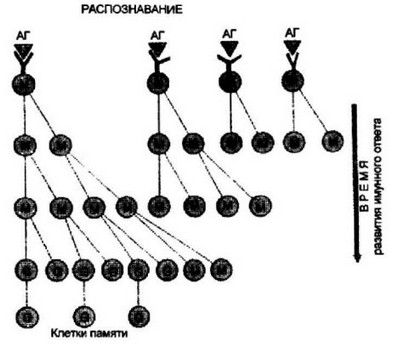
Рис. 9.20. Повышение аффинности антител в динамике развития иммунного ответа.
В реакцию распознавания антигена (АГ) вступают В-клетки нескольких клонов, имеющих близкие, но тем не менее несколько отличающиеся по специфичности антигенраспознающие рецепторы. В силу этого на самых ранних этапах развитии иммунного ответа пул специфических антител будет наиболее гетерогенным. Клон, имеющих антигенраспознающие рецепторы с наибольшим сродством к антигену, будет обладать селективным преимуществом в размножении, что обеспечит ускоренное накопление высокоаффинных антител, именно в этом клоне, количественно большим друнгих клонов, вступят в работу с большей эффективностью механизмы переключения синтеза IgM на IgG и формирования клеток памяти. Схема объясняет как большую аффинность IgG по сравнению с таковой IgM, так и преимущественный синтез высокоаффинных IgG антител при вторичном ответе
генраспознающие иммуноглобулиновые рецепторы, которые несколько отличаются друг от друга по строению антигенсвязываю- щего (активного) центра. В силу этого на самых ранних этапах иммунного ответа пул специфических антител к данному антигенному эпитопу будет наиболее гетерогенным. Клон, имеющий антигенраспознающие рецепторы с наибольшим сродством к антигену, будет обладать селективным преимуществом в размножении за счет большей энергии связи, что и обеспечит ускоренное накопление высокоаффинных антител. Именно в этом клоне вступят в работу механизмы переключения синтеза IgM на IgG и формирования клеток памяти. Схема объясняет как большую аффинность IgG, установленную экспериментально, так и преимущественный синтез высокоаффинных IgG при вторичном ответе, что также имеет экспериментальное подтверждение.
250
Гистологическая картина образования активных анти тело продуцентов. Одной из характерных черт организации лимфоидной ткани является наличие так называемых центров размножения, которые представляют собой место пролиферации, трансформации и селекции В-клеточных клонов (рис. 9.21). В-Лимфоциты, активированные хелперными Т-клетками в тимусзависимой зоне лимфоидной ткани либо сразу дифференцируются в плазматические клетки, продуцирующие ранние, суммарно низкоаффинные антитела, либо перемещаются в первичные фолликулы и образуют там центры размножения. Здесь они, во-первых, подвергаются селекции на наличие высокоаффинных антигенраспознающих иммуноглобулиновых рецепторов и, во-вторых, завершают диффе- ренцировку в плазмоциты, продуцирующие высокоаффинные антитела. Часть В-клеток с высокоаффинными рецепторами трансформируется в клетки памяти.
Первичный фолликул содержит покоящиеся В-клетки, плотно упакованные вокруг специализированных клеточных типов, называемых фолликулярными дендритными клетками. Эти клетки оj - личаются от дендритных клеток, активирующих Т-клетки, по крайней мере по двум признакам. Во-первых, они способны длительное время сохранять нативный антиген на своей поверхности. Во- вторых, данные клетки экспрессируют специализированную молекулу CD23, которая является лигандом для CR2 — компонента корецептора В-клеток CD 19. Взаимодействующие CD23 и CR2 принимают участие в селекции высокоаффинных В-клеток.
Проникшие в первичный фолликул активированные В-клетки начинают интенсивно делиться, образуя властные формы. Они легко определяются микроскопически по увеличенному размеру, обильной цитоплазме, диффузному хроматину. Клетки этого типа называются центробластами. Собственно первичный фолликул, накопивший центробласты, гистологически определяется как центр размножения, или вторичный фолликул. Центробласты дифференцируются в центроциты — это те же В-клетки, но прошедшие активацию Т-лимфоцитами и бластную стадию развития.
Присутствующий на фолликулярных дендритных клетках антиген выступает в качестве фактора отбора центроцитов на наличие у них высокоаффинных антигенраспознающих рецепторов. Этот специфический отбор закрепляется вступлением в реакцию взаимодействия лигандов CD24 и CR2. В-Клетки, не прошедшие отбора, погибают, что микроскопически также определяется по наличию апоптических клеток. Клетки, выдержавшие отбор на аффинность, экспрессируют ген bcl-2+, продукт которого защищает клетку от апоптоза.
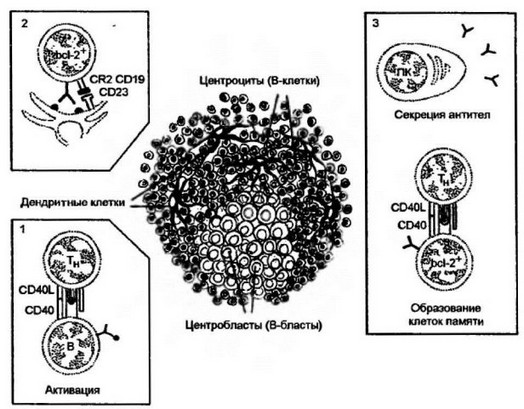
Рис. 9.21, Гистологическая картина функционального созревания В-клеток.
Фолликулы лимфоидной ткани с центрами размножения представляют собой место локализации В-клеток, находящихся на разных стадиях развития. Более светлая часть фолликула — центр размножения — концентрирует бластные формы — центробласты (В-бласты). Центр окружен созревающими или созревшими цен- троцитами (В-клетками). В состав фолликула входят также фолликулярные дедрит- ные клетки, которые отличаются от дендритных клеток, активирующих Т-клетки, двумя признаками:способностью к длительному сохранению нативного антигена на своей поверхности и наличием специализированной молекулы CD23.1. Активация В-клеток (В) хелперными CD4 Т-клетками (Т) в тимусзависимой зоне, прилегающей к фолликулу, приводит in situ к образованию из части активированных клеток ранних плазмоцитов, продуцирующих антитела низкой аффинности. 2. Большая часть активированных В-клеток перемещается в центр фолликула, где начинает активно делиться, образуя бластные формы (центробласты). Микроскопически центр размножения выглядит более светлым. Накопившиеся бласты проходят отбор на повышенную аффинность антиген распознающих рецепторов. Фактором отбора являются дендритные клетки, несущие на своей поверхности чужеродный антиген и экспрессирующие молекулу CD23. Данная молекула выступает при отборе в качестве лиганда, взаимодействующего с СВ2-частью CD 19-рецептора В- клетеж. У прошедших положительный отбор В-клеток происходит дерепрессия гена Ъс1-2+, продукт которого защищает В-клетки от апоптоза. Не прошедшие отбор клетки гибнут на месте, что хорошо видно микроскопически по наличию фигур апоптоза. 3. Прошедшие отбор клетки (центроциты) перемешаются на периферию фолликула, где трансформируются в плазмоциты, секрстирующие антитела высокой аффинности, или образуют клетки памяти при участии хелперных CD4 Т-клеток
Прошедшие сито отбора центроциты перемещаются на периферию центра размножения, где они дифферецируются либо в активно секретирующие антитела плазмоциты, либо в клетки памяти. Что является определяющим в выборе направления диффе- ренцировки на самом заключительном этапе развития, не ясно. Предполагается, что к трансформации в плазмоциты приводит только взаимодействие С1323-лиганда с CR2. В то же время, если зрелые центроциты вступят в контакт с Т-клетками через взаимодействие CD40 с CD40L, то клеточная трансформация приведет к формированию клеток памяти.
Впервые необходимость кооперации Т- и В-лимфоцитов при развитии гуморального иммунного ответа продемонстрировали Клэмен с соавт. в 1966 г. (рис. 9.16). Введение летально облученным мышам клеток только костного мозга (источника В-клеток) или
Рис. 9.16. Взаимодействие между клетками тимуса (источника Т-клеток) и клетками костного мозга (источника В-клеток) при индукции гуморального иммунного ответа (Clamp;man et а)., 1966),
Введение облученным мышам клеток только костного мозга (ККМ) или только тимуса (КТ) не обеспечивает развитие иммунного ответа достаточной силы. В то же время введение смеси этих клеток приводит к интенсивной продукции антител к использованному для иммунизации антигену (эритроцитам барана — ЭБ). Причем ответ при совместной инъекции клеток значительно выше, чем сумма ответов при раздельном введении клеток различного происхождения. Таким образом, кооперация двух типов клеток приводит к синергическому эффекту. Ответ оценивали по количеству бляшкообразующих (антителообразуюших) клеток (БОК) в селезенке
только тимуса (источника Т-клеток) не приводило к развитию гуморального иммунного ответа. Продукция антител находилась на крайне низком уровне. В то же время при одновременной инъекции смеси этих клеток наблюдался сильный иммунный ответ. При одновременном введении различных типов клеток он был значительно выше, чем сумма ответов при раздельной инъекции. Иначе говоря, кооперация Т- и В-клеток дает выраженный синергический эффект. Именно синергизм при клеточном взаимо- 240
действии вызвал наибольший интерес исследователей. Эти опыты определили создание целого направления исследований, задача которого — познание механизмов клеточного взаимодействия при развитии иммунного ответа.
Поверхностный иммуноглобулин (slg) играет двойную роль при трансформации В-клеток в антителопродуцирующие плазмо- циты. Во-первых, подобно антигенраспознающему рецептору Т- клеток он передает сигнал о встрече с антигеном непосредственно внутрь клетки. Во-вторых, служит молекулярным фактором захвата этого антигена для переноса его в цитоплазму клетки. Там антиген разрушается до отдельных пептидов, которые в комплексе с молекулами II класса МНС выносятся на поверхность клетки. Иммуногенный комплекс является объектом распознавания ан- тигенспецифическими хелперными CD4 Т-клетками (Тн2), которые обеспечивают второй сигнал для активации В-клеток (см. рис. 9.9; 3).
Тимусзависимые и тимуснезависимые антигены Для большинства антигенов взаимодействие хелперных Т-клеток с В-лимфо- цитами является обязательным условием развития гуморального иммунного ответа. В силу этого подобные антигены (в основном белки, включающие бактериальные токсины) получили название тимусзависимых антигенов (англ, сокр.: “TD antigens”).
Животные или человек с дисфункцией тимуса не способны к полноценной продукции антител. При подобном иммунодефи- цитком состоянии В-клетки не получают второго сигнала от Т- клеток и переходят в состояние анергии (рис. 9.17).
Хотя для индукции антителогенеза к большинству антигенов требуются зрелые Т-клетки, многие компонентны микробов, такие как бактериальные полисахариды, липополисахарады, высокополимерные белки, могут включать В-клетки без дополнительной помощи со стороны хелперных Т-клеток. Эта категория антигенов получила названгие тимуснезависимых антигенов (англ, сокр.: “TI antigens”).
Тимуснезависимые антигены подразделяются на два класса, которые активируют В-клетки разным способом. TI-1-антигены содержат характеристику, которая проявляется в прямой индукции пролиферации В-клеток. При высокой концентрации антигена в процесс пролиферации и дифференцировки вступят все В-клетки, независимо от специфичности их антигенраспознающих рецепторов. Такие антигены получили название поликлональных активаторов, или В-клеточных митогенов. Примером в данном случае может служить липополисахарид бактерий. Снижение концентрации TI-антигена на несколько порядков приводит к стимуляции только антигенспецифического клона В-клеток. Концентрационный эффект связан с наличием у TI-1 антигенов так называемого
24!
Рис. 9.17. Отлет В-клеток на тнмусзависимый и тимуснсзавксимый антигены.
1. В условиях, когда В-клетки (В) провзаимодейспювали с антигеном (АГ). но не получили помощи со стороны хелперных CD4 Т-клеток (Т), развивается функциональное подавление клетки — анергия. 2. При адекватном включении в ответ хелперных CD4 Т-клеток и формировании в результате двух сигналов для В- клеток (первый — от поверхностного иммуноглобулина (slg), провзаимодейство- вавшего с антигеном (АП, второй — от иммуногенного комплекса на В-клетках) создаются условия для полноценного развития В-клеток в антителопродуиенты. Антигены, которым необходима помощь со стороны Т-клеток, получили название тимуезависимых антигенов. 3. В-клеточный ответ может развиваться без помощи со стороны Т-клеток в тех случаях, когда он формируется на тимуснезависи- мые антигены (TI). Известно два класса таких антигенов. К первому классу (ТЫ) относятся антигены, в чью структуру включат так называемый митогенный участок (МУ), который формирует второй сигнал для В-клеток и тем самым заменяет помощь со стороны Т-клеток. Второй класс — тимуснезависимыс антигены (П-2) — представлен соединениями, в структуре которых имеются повторяющиеся гомологичные эпитопы. Многоточечное соединение этих эпитопов с поверхностными иммуноглобулинами оказывается достаточным для включения В-клстки в процесс пролиферации и дифференцировки до зрелого антителопродуцента митогенного участка, который заменяет сигнал от хелперных Т- клеток. При высоких концентрациях антигена, когда нет конкуренции за специфическое взаимодействие между поверхностным иммуноглобулином и эпитопом TI-1 антигена, наблюдается поликлональный ответ. В условиях, когда концентрация антигена незначительна, преимущество остается за клетками, способными к двойному распознаванию — как митогенного участка, так и антигенного эпитопа. В результате наблюдается клональная экспансия антигенспецифических В-клеток. Для Т1-1 антигена не требуется многократного повторения идентичных эпитопов, так как нали-
чие митогенного участка компенсирует отсутствие многоточечного взаимодействия, что имеет место при активации В-клеток TI - 2-антиге н ам и.
В течение обычного инфекционого процесса концентрация TI-1 -антигенов незначительна. В силу этого будут активироваться только те В-клетки, которые имеют соответствующий антиген- распознающий рецептор, а продуцируемые антитела будут взаимодействовать только с TJ-1 -антигеном. Такой ответ развивается к ряду внеклеточных инфекций, причем он формируется быстрее, чем Т-заьисимый иммунный ответ, так как не требует времени на созревание антигенспецифических хелперных Т-клеток. Скорость ответа сопряжена с несовершенством самого ответа. Он действительно напоминает “скорую помощь” в самом начале развития инфекции. TI-1-антигены, действуя самостоятельно, не способны индуцировать переключение синтеза антител с одного изотипа на другой, не оказывают влияния на повышение аффинности антител, не создают клеток памяти. Все эти процессы требуют хелперных Т-клеток.
Второй класс тимуснезависимых антигенов представлен молекулами, имеющими часто повторяющиеся идентичные эпитопы. К этому классу относятся полисахариды бактериальной стенки, сильно полимеризованные высокомолекулярные белки (например, гемоцианин). Механизм активации В-клеток связан с перекрестным сцеплением между повторяющимися антигенными эпитопами и поверхностным иммуноглобулином. Является ли ответ к TI-i-антигенам полностью независимым от хелперных Т- клеток, не ясно. С одной стороны, гуморальный иммунный ответ регистрируется у дефицитных по Т-клеткам мышей nude. С другой — удаление из культуры in vitro всех Т-клеток приводит к подавлению ответа на TI-2-антиген. Введение незначительного количества Т-клеток иммунодефицитным мышам значительно усиливает иммунный ответ к данным антигенам. Вероятно, наиболее ранний ответ в виде синтеза IgM формируется без помощи Т-клеток. Более поздний гуморальный ответ, характеризующийся доминантным синтезом IgG, требует включения хелперных Т-клеток.
Сцепленное распознавание. В условиях конкретной вирусной или бактериальной инфекции В- и Т-клетки должны распознать тот же самый комплексный антиген, но не обязательно те же самые эпитопы этого антигена. Понимание того факта, что В- и Т- клетки могут реагировать с разными эпитопами и что при этом помощь со стороны хелперных Т-клеток проявляется в полной мере, пришло из опытов по индукции гуморального иммуного ответа у животных, иммунизированных комплексом гаптенгноси- тель (рис. 9.18). Мышей иммунизировали конъюгатом NIP-OA (N1P — гаптен 4-гидрокси-З-йод- 5-нитрофенилуксусная кислота; ОА
243
Таблица 9.4
Сравнительная характеристика различных классов антигенов, инициирующих ^моральный иммунный ответ
|
Свойства |
Тимусзависимые (TI) антигены: токсин дифтерии, вирусный гемагглютинин, очищенный белковый дериват (PPD) Mycobacterium tuberculosis |
Ти мусн ела виси- мыс-1 (ТЫ) антигены: бактериальный липо- пслисахарид Brucella abortus |
Тимуснезависи- мыс-2 (TI-2) антигены: полисахарид пневмококков, полимерилованный флагеллин сальмонелла, по- ливинилпирол- лидон, фиколл, конъюгированный с гаптеном |
|
Образование антител у нормальных животных |
да |
да |
да |
|
Образование антител у дефицитных по Т-клеткам животных |
нет |
ла |
да |
|
Примированис Т-клсток |
да |
нет |
нет |
|
Поликлональная активации В-кле- теж |
нет |
да |
нет |
|
Требование к повтори юшимся эпитопам |
нет |
нет |
да |
— овальбумин кур, использованный в качестве носителя для гап- тена). При переносе клеток селезенки примированных мышей в организм сингенного облученного хозяина развивается анти-NlP- ответ только после введения того же антигена NIP-OA. Продукция антител к гаптену отсутствовала, если в качестве антигена использовали NIP, конъюгированный с другим носителем, например с бычьим сывороточным альбумином (BSA) или сывороточным альбумином человека (HSA). Отсутствовал ответ и в тех случаях, когда перед введением реципиенту клеток селезенки освобождали от Т-клегок, используя анти-Thy-l -сыворотку. Эта первая часть опытов демонстрировала, во-первых, необходимость примированных Т-клеток для развития гуморального иммунного ответа, и во-вторых, распознавание разных эпитопов NIP-OA конъюгата. Если бы Т-клетки распознавали тот же гаптен, то его введение на ином носителе не отразилось бы на силе гуморального ответа. В последующих вариантах этих опытов проводили адаптивный перенос примированных клеток селезенки от мышей иммунизированных как NIP-OA, так и только посторонним носителем — BSA. В последнем случае иммунный ответ развивался и к NIP-OA, и к NIP- 244
BSA. Удаление Т-клеток из селезенки мышей, иммунизированных BSA, отменяло и продукцию антител к гаптену. Ясно, что имму- ногенность конъюгата связана с возможностью Т-клеток взаимодействовать с носителем. При этом антигенные свойства носителя не имеют значения для специфичности гуморального иммунного ответа.
При реальной бактериальной или вирусной инфекции, как и в эксперименте, Т- и В- клетки распознают разные эпитопы одного и того же комплексного в антигенном отношении патогена. Это явление распознавания В- и Т-клетками разных эпитопов одного и того же антигена получило название сцепленного распознавания. На примере вирусной инфекции цепь событий выглядит следующим образом. Поверхностные белки вирусных частиц, взаимодействуя с предетерминированными антигенраспознающими рецепторами (slg) В-клеток, оказываются поглощенными этими В-клетками. В результате внутриклеточной переработки на клеточной поверхности оказываются пептиды вирусных белков в комплексе с молекулами II класса МНС. Комплекс образуется с пептидами не только внешних вирусных белков, но и внутренних, недоступных для поверхностных иммуноглобулинов В-клеток. Таким образом, В-клетка оказывается подготовленной к встрече со зрелыми хелперными Т-клетками. Т-хелперы проходят свой путь при- мирования, взаимодействуя с тем или иным типом антигенпре- зентирующих клеток. Среди Т-клеток будут те, которые способны реагировать с иммуногенными комплексами, включающими пептиды как внешних, так и внутренних белков. Таким образом помощь В-клеткам усиливается способностью Т-клеток распознавать множество пептидных вирусных фрагментов, но при этом специфичность секретируемых В-ютеткой антител будет только к поверхностному белку вируса, так как первоначальная встреча В- лимфоцитов была именно с поверхностными эпитопами. Образовавшиеся вирусспецифические антитела обладают как нейтрализующей, так и опсонизирующей активностью, т.е. с одной стороны, препятствуют взаимодействию вируса с клеткой-мишенью, а с другой — усиливают контакт с фагоцитирующими клетками через Fc-фрагмент антител.
Мембранные и секреторные активаторы В-клеток. Зрелые хел- перные CD4 Т-клетки, распознавшие комплекс антигенного пептида с молекулами II класса МНС, активируют В-клетки к пролиферации и дифференцировке в зрелые антителопродуценты — плазмоциты. Хелперные Т-клетки после взаимодействия с В-лим- фоцитами начинают экспрессировать мембраносвязанные и секреторные формы молекулярных активаторов (рис. 9.19). Наиболее важными эффекторными молекулами хелперных Т-клеток на ранней стадии активации В-клеток является лиганд CD40, относя- 246
Рис. 9.19. Активация В-клеток к пролиферации и дифференцировке в плазмоци-
ты.
В-клетки (В), экспрессирующие комплекс антигенный пептид:молекула II класса МНС, взаимодействуют с антигснспе пифически ми хелпсрными Т-клетка- ми ГГц). При этом формируются дополнительные сигналы, необходимые В-клетке для выхода в дифференцировку: первый сигнал — от взаимодействия CD40 В- клеток с лигандом CD40L Т-клетсж и второй сигнал — от секретируемого Т- хелперамицитокина интсрлейкин-4 (ИЛ-4). Два дополнительных цитокина — интерлейкин-5 (ИЛ-5) и интерлейкин-6 (ИЛ-6), секрета русмы е Т-клеткам и, вносят свой вклад в процесс дифференцировки В-клеток на заключительных этапах развития
щийся к семейству ФНО-цитокинов. Этот лиганд связывается с поверхностной молекулой В-клеток CD40 — аналогом рецептора к ФНО-ct на макрофагах. Вторым стимулирующим В-клетки фактором является интерлейкин-4 (ИЛ-4), секреция которого хелпер- ными Т-клетками начинается после их контакта с В-лимфоцита- ми. Как и в случае с инфицированными макрофагами, требующими для своей активации двух молекулярных сигналов, синтезируемых Т-клетками воспаления (поверхностного ФНО-а и секрети- руемого цитокина ИНФ-у), В-клетки также активируются двумя стимуляторами — поверхностным СО40-лигандом и секретируе- мым ИЛ-4. Однако если активация инфицированных макрофагов приводит к быстрому эффекторному действию — разрушению внутриклеточных патогенов, то начальный этап активации наивных В- клеток связан с рядом подготовительных процессов, и в первую очередь с их клональной экспансией — накоплением антигенспе- цифического клона, который после завершения пролиферации переходит к дифференцировке в плазматические клетки, секрети- рующие антитела заданной специфичности. Два дополнительных цитокина — ИЛ-5 и ИЛ-6, секретируемые хелпервыми Т-клетками, вносят свой вклад в процесс дифференцировки В-клеток на заключительном этапе процесса В-клсточной активации.
Переключение синтеза изотипов. По мере развития гуморального иммунного ответа синтез антител переключается с одного изотипа на другой при сохранении их исходной специфичности. В самом начале иммунного ответа доминирующим изотипом является IgM. В очень незначительном количестве представлен IgD. Позднее, по мерс развития антителогенеза происходит замена синтеза IgM на синтез IgG, IgA и IgE. Эти изменения в преимущественном синтезе того или иного изотипа не наблюдаются у индивидуумов с дефектом экспрессии СО40-лиганда, принимающего участие в контакном взаимодействии хелперных Т-клеток с В-лимфо- иитами. При подобной форме иммунодефицита наблюдается необычно высокое содержание IgM в сыворотке крови на фоне отсутствия продукции иммуноглобулинов иных изотипов. Активная продукция IgM при недостатке Т-клеточной помощи может быть связана с ответом В-клеток на тимуснезависимые антигены. В то же время эти клинические наблюдения дополнительно демонстрируют, что для сбалансированной продукции различных классов антител необходимо включение в ответ хелперных Т-клеток.
Большинство из того, что известно о регуляции переключения изотипов, пришло из экспериментов, проведенных с мышиными В-клетками, которые стимулировались очищенными цитокинами in vitro. Эти эксперименты показали, что различные цитокины преимущественно индуцируют переключение синтеза “своих” изотипов. Так, ИЛ-4 индуцирует переключение синтеза IgM на IgGI и lg, в то время как ФНО-р — на IgG2b и IgA. Другой цитокин — ИЛ-5, секретируемый хелперными CD4 Т-клетками, усиливает продукцию IgA, когда переключение в геноме для синтеза данного изотипа уже произошло. Хотя CD4 Т-клетки воспаления являются плохими регуляторами синтеза антител, они принимают участие в переключении изотипов посредством секрети- руемого ими ИНФ-у. Этот цитокин оказывает преимущественное влияние на индукцию синтеза IgG3 и IgG2a.
Данные об участии различных цитокинов в продукции антител разных изотипов суммированы в табл. 9.5.
Таким образом, участие хелперных Т-клеток в гуморальном иммунном ответе имеет двойное проявление: во-первых, они инициируют пролиферацию и дифференцировку распознавших антиген В-клеток и, во-вторых, оказывают прямое действие на переключение синтеза антител с одного изотипа на другой.
Повышение аффинности антител в процессе формкрования.им- муиного ответа. По мере развития иммунного ответа происходит не только переключение синтеза антител с одного изотипа на другой, но и повышается аффинность (сродство) этих антител к тому антигенному эпитопу, который вызвал их образование. Константа диссоциации “ранних” антител на два-три порядка выше, чем у
Таблица 9.5
Влияние цитокинов на экспрессию различных изотипов
иммуноглобулинов
“поздних” антител. Кроме того, поданным изоэлектофокусирова- ния известно, что на раннем этапе иммунного ответа гетерогенность антител одной специфичности выше, чем на поздних этапах процесса.
Одна из гипотез, пытающихся объяснить это явление, строится на представлении о гипермутабелыюсти генов, контролирующих вариабельные домены иммуноглобулинов. Известно, что мутационный уровень для всех соматических клеток составляет замену одной пары оснований на 1012 пар при одном клеточном делении. По гипермутационпой теории частота замен представляется величиной, равной одной замене на 103 пар оснований. При этом гипермутабельность не затрагивает гены, контролирующие константные регионы. Первое и основное возражение в отношении данной гипотезы заключается в том, что представление о выборочной повышенной мутабельности каких-то участков ДНК (в данном случае V-генов) кажется умозрительным и не укладывается в современные концепции молекулярной генетики. Непонятно, зачем строить искусственные конструкции, когда уровень исходной генетически детерминированной вариабельности 2,4* 10* разных по специфичности иммуноглобулинов более чем достаточен для взаимодействия с самыми различными антигенными эпитопами; среди них будут и такие, взаимодействие с которыми осуществляется с наибольшей аффинностью.
Вопрос разрешается достаточно просто на основании представлений о существовании сыекции антигенспецифических В- ююнов. На рис. 9.20 показан один из возможных механизмов повышения аффинности антител в процессе развития иммунного ответа зрелыми В-клетками. В реакцию распознавания антигенного эпитопа вступают клетки нескольких клонов, имеющих анти-
Рис. 9.20. Повышение аффинности антител в динамике развития иммунного ответа.
В реакцию распознавания антигена (АГ) вступают В-клетки нескольких клонов, имеющих близкие, но тем не менее несколько отличающиеся по специфичности антигенраспознающие рецепторы. В силу этого на самых ранних этапах развитии иммунного ответа пул специфических антител будет наиболее гетерогенным. Клон, имеющих антигенраспознающие рецепторы с наибольшим сродством к антигену, будет обладать селективным преимуществом в размножении, что обеспечит ускоренное накопление высокоаффинных антител, именно в этом клоне, количественно большим друнгих клонов, вступят в работу с большей эффективностью механизмы переключения синтеза IgM на IgG и формирования клеток памяти. Схема объясняет как большую аффинность IgG по сравнению с таковой IgM, так и преимущественный синтез высокоаффинных IgG антител при вторичном ответе
генраспознающие иммуноглобулиновые рецепторы, которые несколько отличаются друг от друга по строению антигенсвязываю- щего (активного) центра. В силу этого на самых ранних этапах иммунного ответа пул специфических антител к данному антигенному эпитопу будет наиболее гетерогенным. Клон, имеющий антигенраспознающие рецепторы с наибольшим сродством к антигену, будет обладать селективным преимуществом в размножении за счет большей энергии связи, что и обеспечит ускоренное накопление высокоаффинных антител. Именно в этом клоне вступят в работу механизмы переключения синтеза IgM на IgG и формирования клеток памяти. Схема объясняет как большую аффинность IgG, установленную экспериментально, так и преимущественный синтез высокоаффинных IgG при вторичном ответе, что также имеет экспериментальное подтверждение.
250
Гистологическая картина образования активных анти тело продуцентов. Одной из характерных черт организации лимфоидной ткани является наличие так называемых центров размножения, которые представляют собой место пролиферации, трансформации и селекции В-клеточных клонов (рис. 9.21). В-Лимфоциты, активированные хелперными Т-клетками в тимусзависимой зоне лимфоидной ткани либо сразу дифференцируются в плазматические клетки, продуцирующие ранние, суммарно низкоаффинные антитела, либо перемещаются в первичные фолликулы и образуют там центры размножения. Здесь они, во-первых, подвергаются селекции на наличие высокоаффинных антигенраспознающих иммуноглобулиновых рецепторов и, во-вторых, завершают диффе- ренцировку в плазмоциты, продуцирующие высокоаффинные антитела. Часть В-клеток с высокоаффинными рецепторами трансформируется в клетки памяти.
Первичный фолликул содержит покоящиеся В-клетки, плотно упакованные вокруг специализированных клеточных типов, называемых фолликулярными дендритными клетками. Эти клетки оj - личаются от дендритных клеток, активирующих Т-клетки, по крайней мере по двум признакам. Во-первых, они способны длительное время сохранять нативный антиген на своей поверхности. Во- вторых, данные клетки экспрессируют специализированную молекулу CD23, которая является лигандом для CR2 — компонента корецептора В-клеток CD 19. Взаимодействующие CD23 и CR2 принимают участие в селекции высокоаффинных В-клеток.
Проникшие в первичный фолликул активированные В-клетки начинают интенсивно делиться, образуя властные формы. Они легко определяются микроскопически по увеличенному размеру, обильной цитоплазме, диффузному хроматину. Клетки этого типа называются центробластами. Собственно первичный фолликул, накопивший центробласты, гистологически определяется как центр размножения, или вторичный фолликул. Центробласты дифференцируются в центроциты — это те же В-клетки, но прошедшие активацию Т-лимфоцитами и бластную стадию развития.
Присутствующий на фолликулярных дендритных клетках антиген выступает в качестве фактора отбора центроцитов на наличие у них высокоаффинных антигенраспознающих рецепторов. Этот специфический отбор закрепляется вступлением в реакцию взаимодействия лигандов CD24 и CR2. В-Клетки, не прошедшие отбора, погибают, что микроскопически также определяется по наличию апоптических клеток. Клетки, выдержавшие отбор на аффинность, экспрессируют ген bcl-2+, продукт которого защищает клетку от апоптоза.
Рис. 9.21, Гистологическая картина функционального созревания В-клеток.
Фолликулы лимфоидной ткани с центрами размножения представляют собой место локализации В-клеток, находящихся на разных стадиях развития. Более светлая часть фолликула — центр размножения — концентрирует бластные формы — центробласты (В-бласты). Центр окружен созревающими или созревшими цен- троцитами (В-клетками). В состав фолликула входят также фолликулярные дедрит- ные клетки, которые отличаются от дендритных клеток, активирующих Т-клетки, двумя признаками:способностью к длительному сохранению нативного антигена на своей поверхности и наличием специализированной молекулы CD23.1. Активация В-клеток (В) хелперными CD4 Т-клетками (Т) в тимусзависимой зоне, прилегающей к фолликулу, приводит in situ к образованию из части активированных клеток ранних плазмоцитов, продуцирующих антитела низкой аффинности. 2. Большая часть активированных В-клеток перемещается в центр фолликула, где начинает активно делиться, образуя бластные формы (центробласты). Микроскопически центр размножения выглядит более светлым. Накопившиеся бласты проходят отбор на повышенную аффинность антиген распознающих рецепторов. Фактором отбора являются дендритные клетки, несущие на своей поверхности чужеродный антиген и экспрессирующие молекулу CD23. Данная молекула выступает при отборе в качестве лиганда, взаимодействующего с СВ2-частью CD 19-рецептора В- клетеж. У прошедших положительный отбор В-клеток происходит дерепрессия гена Ъс1-2+, продукт которого защищает В-клетки от апоптоза. Не прошедшие отбор клетки гибнут на месте, что хорошо видно микроскопически по наличию фигур апоптоза. 3. Прошедшие отбор клетки (центроциты) перемешаются на периферию фолликула, где трансформируются в плазмоциты, секрстирующие антитела высокой аффинности, или образуют клетки памяти при участии хелперных CD4 Т-клеток
Прошедшие сито отбора центроциты перемещаются на периферию центра размножения, где они дифферецируются либо в активно секретирующие антитела плазмоциты, либо в клетки памяти. Что является определяющим в выборе направления диффе- ренцировки на самом заключительном этапе развития, не ясно. Предполагается, что к трансформации в плазмоциты приводит только взаимодействие С1323-лиганда с CR2. В то же время, если зрелые центроциты вступят в контакт с Т-клетками через взаимодействие CD40 с CD40L, то клеточная трансформация приведет к формированию клеток памяти.