
Мембранный иммуноглобулин, провзаимодействовавший с антигеном, не может сам по себе активировать ответ В-клеток. Связано это с тем, что его цитоплазматический хвост слишком короток. Он содержит всего несколько аминокислотных остатков, не способных вступать в реакционные отношения с внутриклеточными компонентами. Для переноса сигнала с антигенраспознаю- щего рецептора внутрь клетки имеются специальные, низкомолекулярные белки с достаточно длинным хвостовым участком, находящиеся на клеточной поверхности в непосредственной близости от мембранного иммуноглобулина. Всего таких полипептидов два — Iga и Ig|3. Белок Iga специфичен по отношению к тому или иному изотипу иммуноглобулина. В то же время Igp является общим для всех изотипов (рис. 3.3).
Помимо передачи сигнала эти белки выполняют роль "извозчиков”, способствуя выходу мембранного иммуноглобулина на поверхность клетки. В отсутствие этих белков иммуноглобулины остаются во внутриклеточном компартменте.
Активирование В-клеток через взаимодействие антигена с рецептором может происходить как самостоятельно, так и с помощью цитокинов, продуцируемых Т-хелперами и макрофагами. В
(рецепторной) и секреторной форм иммуноглобулина.

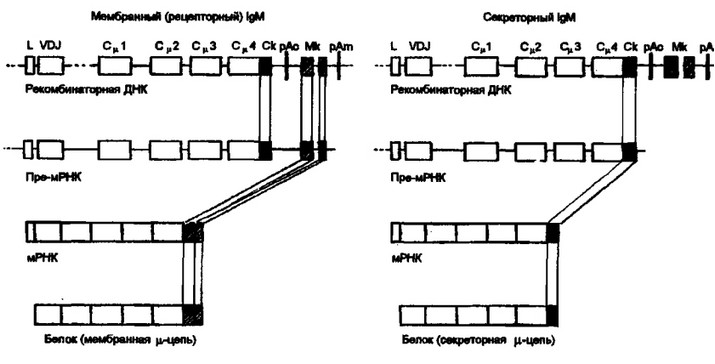 При образовании мембранной или секреторной форм иммуноглобулина процесс реорганизации генома характеризуется следующими особеностями. Рекомбинантная ДНК содержит последовательность нуклеотидов от лидерного участка (L) до терминальной последовательности полиаденина (сокр.: рАш — полиаденин мембранного пути развития) включительно. В реорганизованный локус входят экзон Ск (секреторный компонент) и два экзона Мк (мембранный компонент), один из которых контролирует трансмембранную последовательность, а второй — цитоплазматическую последовательность аминокислотных остатков рецепторного иммуноглобулина. В тех случаях, когда образующийся первичный транскрипт (пре-мРНК) включает экзоны Мк, синтезируется ц-цепь мембранного иммуноглобулина. При этом Ск и рАс (полиаденин секреторного пути реорганизации) подвергаются делении при сплайсинге пре-мРНК. Если первичный транскрипт не включает последовательность Мк и рАт, но сохраняет Ск и рАс, то в результате последующих преобразований синтезируется ц-цепь секреторного иммуноглобулина
При образовании мембранной или секреторной форм иммуноглобулина процесс реорганизации генома характеризуется следующими особеностями. Рекомбинантная ДНК содержит последовательность нуклеотидов от лидерного участка (L) до терминальной последовательности полиаденина (сокр.: рАш — полиаденин мембранного пути развития) включительно. В реорганизованный локус входят экзон Ск (секреторный компонент) и два экзона Мк (мембранный компонент), один из которых контролирует трансмембранную последовательность, а второй — цитоплазматическую последовательность аминокислотных остатков рецепторного иммуноглобулина. В тех случаях, когда образующийся первичный транскрипт (пре-мРНК) включает экзоны Мк, синтезируется ц-цепь мембранного иммуноглобулина. При этом Ск и рАс (полиаденин секреторного пути реорганизации) подвергаются делении при сплайсинге пре-мРНК. Если первичный транскрипт не включает последовательность Мк и рАт, но сохраняет Ск и рАс, то в результате последующих преобразований синтезируется ц-цепь секреторного иммуноглобулина
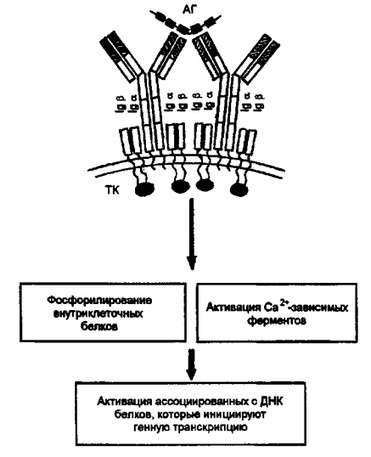
Рис. 3.3. Активация В-клеток ори взаимодействии с антигеном.
Тимус независимый антиген (АГ), взаимодействуя с мембранным (рецепторным) иммуноглобулином, приводит к его агрегации. В результате Iga и Igp, входящие в антигенраспознающий комплекс, взаимодействуют в цитоплазме с тироаинкиназами (ТК), которые в свою очередь обеспечивают ряд внутриклеточных биохимических процессов, инициирующих транскрипцию иммуноглобулиновых генов
первом случае инициация ответа обеспечивается агрегацией рецепторов при их перекрестном связывании соответствующим лигандом. Перекрестное связывание рецепторов происходит в тех случаях, когда антиген обладает повторяющимися, идентичными эпитопами. Для реализации ответа таким антигенам не нужна помощь со стороны Т-хелперов. Отсюда их название — тимуснезави- симые антигены. К ним относятся, в частности, полисахариды бактерий.
Образование агрегатов мембранных иммуноглобулинов под влиянием антигена (в англ, литературе — “cupformation”) приводит к взаимодействию различных тирозинкиназ с Iga и Ig{3. Активированные киназы провоцируют каскад реакций, в результате которых в клетке накапливаются Са2+-зависимые ферменты и происходит фосфорилирование внутриклеточных белков. Именно они
на заключительном этапе формирования ответа В-клеток активируют связанные с ДНК белки и таким образом инициируют транскрипцию специфических генов.