
Организация генов, кодирующих а- и (3-цепи ТКР, в основном гомологична той, которая известна для легких и тяжелых цепей иммуноглобулинов. V-домен a-цепи, подобно легкой цепи иммуноглобулинов, контролируется только V- и J-генными сегментами. В то же время образование V-домена (3-цепи, как и тяжелой цепи иммуноглобулинов, обеспечено полным набором V-, D-, J- генных сегментов (рис. 3.14).
В геноме Т-клеток имеется более 100 V-генов для а-цепи
ТКР, что в два с половиной раза меньше того количества, которое известно для легких цепей иммуноглобулинов. Каждый такой ген включает два экзона — один для лидерной (L) последовательности, отсутствующей у зрелой a-цепи, но представленной у этой цепи в момент ее транспорта из эндоплазматического ретикулума к клеточной поверхности, и второй — для кодирования собственно V-домена ТКР. J-Генных сегментов для a-цепи значительно больше, чем для легкой цепи иммуноглобулинов (50 против 4). Константная область a-цепи контролируется С-геном, включающим отдельные экзоны для С-домена, шарнира и один общий экзон — для трансмембранной и хвостовой частей молекулы.
Количество V-генов для (3-цепи равно 30. Кроме того, имеется два кластера DJC. Каждый кластер включает один D- и шесть J-генных сегментов. Функциональные различия между кластерами неизвестны. С-ген для константной области |3-цепи включает четыре экзона для константного, шарнирного, трансмембранного и хвостового участков полипептида.
Процессы рекомбинации, транскрипции, сплайсинга и трансляции генетического материала для а- и (3-цепей при образовании ТКР в Т-клетках аналогичны тем, которые обеспечивают синтез иммуноглобулинов в В-клетках.
Так же как в случае с иммуноглобулинами и иммуноглобулиновыми рецепторами, вариабельность ТКР зависит от случайного взаимодействия генных сегментов в процессе рекомбинации генетического материала, кодирующего V-домены: VJ — для и- цепей и VDJ -для (3-цепей. Расчет вариабельности V-доменов ТКР, который проводится так же, как и для иммуноглобулинов (см. гл.
- , показывает крайне высокий уровень разнообразия этих анти-
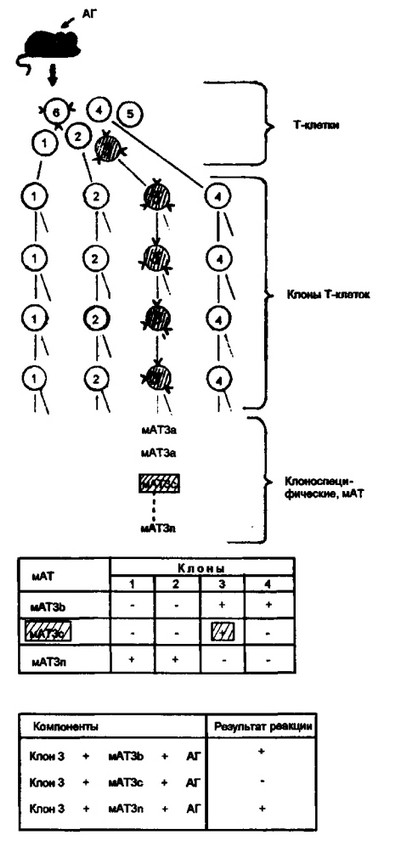
Рис. 3.12. Получение моноклональных антител, специфичных к Т-клеточному антигенраспознающему'рецептору.
На первом этапе работы от мышей, иммунизированных опеределенным антигеном (АГ), получали суммарную, недифференцированную популяцию Т-кле- ток, содержащую самые различные клоны (на рис. цифры 1 -6). Второй этап состоял в выделении отдельных клонов Т-клеток, среди которых были и специфичные к использованному антигену (на рис. в качестве примера приведено четыре клона, один из которых — клон 3, — специфически реагирует с антигеном). Третий этап работы включал получение моноклональных антител (мАТ) к антигенреактивно- му клону. Задача этого этапа — получение моноклональных антител, способных реагировать только с клоном, использованным для иммунизации. В то же время перекрестная реакция мАТ говорит об общей специфике антигенреактивного клона и непримированных клонов (верхняя таблица). Отсутствие перекрестной реактивности мАТ указывает на наличие у положительно реагирующего примирован- ного клона особой специфичности — предположительно, антигенраспознающего рецептора. Подтверждением подобного предположения является реакция задержки взаимодействия мАТ с соответствующим клоном в присутствии использованного антигена (нижняя таблица). Получение мАТ к антигенраспознающему рецептору Т-клеток создало условия для его полноценного изучения
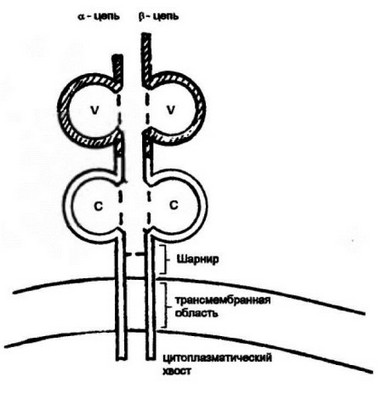
Рис. 3.13. Структура янтнгецрвсооamp;яощсго рецсширя Т-влето*.
Т-клеточный антигенраспознающий рецептор (ТКР) представляет собой гетеродимер, составленный из а- и p-цепей. Каждая цепь включает два домена: вариабельный (V) и константный (С). Взаимодействующие Va- и Ур домены формируют антигенраспознающий участок ТКР. Помимо основных V- и С-доменов в структуре ТКР имеется шарнирная область с цистеиновым остатком, образующим ковалентную связь между а- и p-цепями, а также трансмембранный и короткий хвостовой участки
генраспознающих структур (табл. 3.2). При общем сходстве организации и рекомбинации генетического материала для иммуноглобулинов и ТКР следует отметить и некоторые особенности в контроле специфичности этих молекул.
Иммуноглобулины и иммуноглобулиновые рецепторы 6-клеток распознают нативные антигенные эпитопы. В связи с этим отдельные участки антигенраспознающего центра имеют равные шансы на изменчивость. Ситуация с ТКР несколько иная, поскольку этот рецептор распознает комплекс антигенного пептида с молекулами МНС.
Разнообразие ТКР связано в значительной степени с третьей петлей V-домен а, формируемой третьим гипервариабельным участком — CDR3 (сокр. от англ, “complementarity determining region”). При образовании антигенсвязывающего центра V-доме- нами а- и 0-цепей CDR3 оказываются во внутренней части этого центра. Первая и вторая петли (CDR1 и CDR2, соответственно) занимают периферию центра. В таком конформационном построе
на
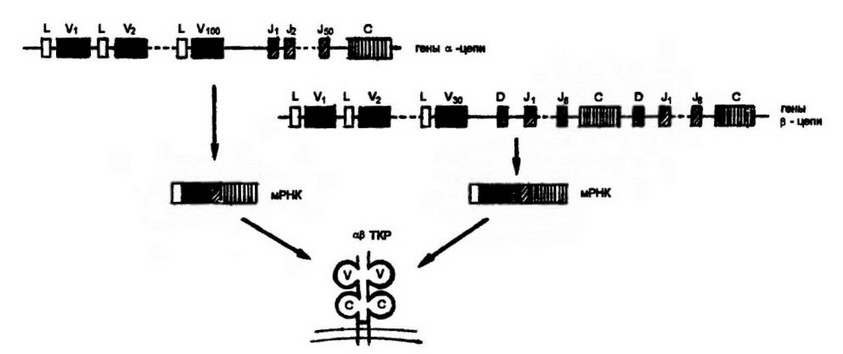
Рис. 3.14. Ортшнзацп геоон, контролирующих а- и р-цеш Т-исшюго рецытгоц.
Принцип реорганизации генных сегментов, контролирующих а- и р-цспи ТКР, таг же, что и для иммуноглобулинов. Отличие состоит в том, что локус для (3-цепи имеет два идентичных кластера. В чем функциональное значение такого дублирования, неизвестно
Таблица 3.2
Вариабельность Т-клеточных рецепторов в сравнении с
иммуноглобулиновыми рецепторами и иммуноглобулинами
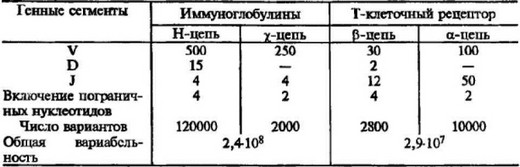
нии имеется вполне определенный биологический смысл, связанный с адаптацией ТКР к той форме антигена, с которой он взаимодействует. Как уже отмечалось, антигенные пептиды заполняют пространство (щель), образованное а-спиральными структурами молекул МНС и таким образом оказываются в середине антигенного комплекса пептидгМНС. Подобный комплекс характеризуется огромным множеством антигенных специфичностей, связанных с пептидами, и ограниченным разнообразием, свойственным молекулам МНС. В связи с подобной организацией иммуногенного комплекса следует ожидать повышенную изменчивость CDR3 и меньшую изменчивость CDR1 и CDR2. Изучение генетической организации генов для ТКР подтверждает подобную точку зрения. Так, ТКР имеет значительно меньшее по сравнению с иммуноглобулинами количество V-генов, определяющих специфичность CDR1 и CDR2, но, при этом увеличенное число J-сегментов, принимающих участие в кодировании CDR3 (см. табл. 3.2). На рис. 3.15 представлена упрощенная схема, иллюстрирующая взаимодействие СОЮ с пептидом и CDR1 и CDR2 — с а-спиральной последовательностью молекул МНС.