
ТКР, как и мембранный, антигенраспознающий иммуноглобулин В-клеток, имеет очень короткий цитоплазматический хвост. В связи с этим сигнал от взаимодействия ТКР с комплексом пептид: молекулы МНС не может быть передан внутрь клетки. Трансмиссивную функцию выполняют инвариантные, низкомолекулярные, ассоциированные с ТКР белки, которые получили общее название — CD3. Комплекс CD3 включает пять белков: три белка CD3T, CD36 и CD3e, представленных на клеточной поверхности и имеющих определенную гомологию с иммуноглобулинами, и два
104
Рис. 3.15- Принцип cipjLijrpiux отношений между птервgt;ркАбельиымн участками Т-клеточиого аэтнгенрше- познающего рецептора н комплексом пептцд:молекулы главного комплекса гнстосоаместемосп!.
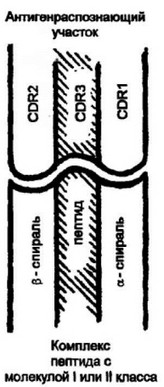 Гипервариабельные участки ТКР конформаци- онно организованы так, что наиболее вариабельный регион (CDR3) локализован в середине антигенрас- познающего участка. При этом распознаваемый тид также находится в середине комплекса между а- спиральными последовательностями молекул МНС. Подобная конформационная особенность обеспечивает наиболее эффективное распознавание пептида в комплексе
Гипервариабельные участки ТКР конформаци- онно организованы так, что наиболее вариабельный регион (CDR3) локализован в середине антигенрас- познающего участка. При этом распознаваемый тид также находится в середине комплекса между а- спиральными последовательностями молекул МНС. Подобная конформационная особенность обеспечивает наиболее эффективное распознавание пептида в комплексе
цитоплазматических белка —
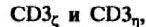 не имеющих такой
не имеющих такой
гомологии (рис. 3.16).
Белки, гомологичные иммуноглобулинам, экспрессируются на клеточной поверхности в виде гетеродимеров
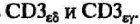 Их связь с ТКР осуществляется посредством электростатического притяжения. Отрицательно заряженные трансмембранные участки цепей CD3 взаимодействуют с несущими положительный суммарный заряд трансмембранными участками ТКР. Наличие длинного хвоста позволяет им взаимодействовать с цитоплазматическими белками-трансдукторами после получения антигенного сигнала.
Их связь с ТКР осуществляется посредством электростатического притяжения. Отрицательно заряженные трансмембранные участки цепей CD3 взаимодействуют с несущими положительный суммарный заряд трансмембранными участками ТКР. Наличие длинного хвоста позволяет им взаимодействовать с цитоплазматическими белками-трансдукторами после получения антигенного сигнала.
Два других полипептида —
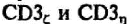 - также входят в
- также входят в
состав комплекса в виде димеров _
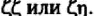 _. Около 80% ТКР ассо
_. Около 80% ТКР ассо
циировано с гомодимером и только 20% - с гетершимером. Функциональные различия между ними неизвестны. Основной домен этих белков в отличие от других CD3-белков находится в цитоплазме. Именно головная, а не хвостовая часть
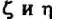 взаимодей
взаимодей
ствует в цитоплазме с белками-трансдукторами.
Помимо сигналпередающей функции, белки CD3 ответственны за транспорт ТКР к клеточной поверхности. У мутантных клеток, в которых отсутствует синтез
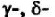 или
или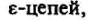 экспрессия
экспрессия
ТКР полностью подавлена, хотя внутриклеточный синтез этих рецепторов не нарушен. При мутациях гена
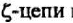 выход ТКР иа
выход ТКР иа
клеточную поверхность происходит в меньшей степени по сравнению с нормой.
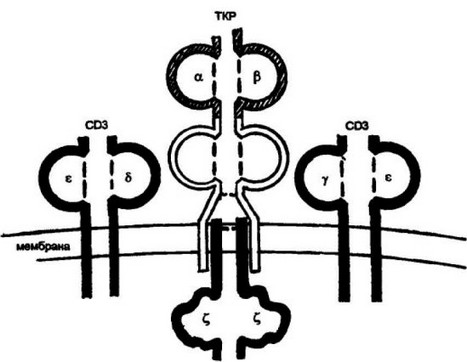
Рис. 3.16. Строение Т-клеточного ашнгещмаюзшиощего комплекс*.
Комплекс включает Т-клеточный антигенраспоэншощий рецептор (ТКР) и пять однодоменных инвариантных белков: у,6, г находятся на поверхности клетки; ?иг| погружены в цитоплазму; т\ не отмечен на рисунке; у,8,в, имеющие удлиненный цитоплазматический хвост, служат для передачи сигнала внутрь клетки после взаимодействия ТКР с антигеном
Как трансмиссивная, так и транспортная функции СБЗ-бел- ков гомологична той, которая характерна для Iga- и Igp-белков иммуноглобулинового, антигенраспознающего комплекса.
В активации Т-клеток, распознавших антиген, также принимают участие CD4 и CD8 — маркеры дифференцировки Т-клеток. Как уже отмечалось, первый из них является маркером CD4 Т- клеток, второй — цитотоксических Т-лимфоцитов (CD8 Т-клеток). Долгое время функция этих белков оставалась неизвестной. Оказалось, что они принимают самое непосредственное участие в процессе взаимодействия ТКР с соответствующим лигандом в качестве корецептора.
CD4 представляет собой одноцепочечную молекулу, состоящую из четырех иммуноглобул и нподобных доменов (рис. 3.17). Домены D| и D2, а также D3 и D4 образуют между собой парные, плотноупакованные, жесткие структуры. Эти пары соединены гибким шарнирным участком. Хвостовая часть молекулы CD4 имеет достаточную длину для взаимодействия с цитоплазматическими белками-трансдукторами. На клеточной поверхности ТКР и CD4 представлены независимо друг от друга. Их встреча происходит в процессе формирования ответа на антиген. После распознавания 106
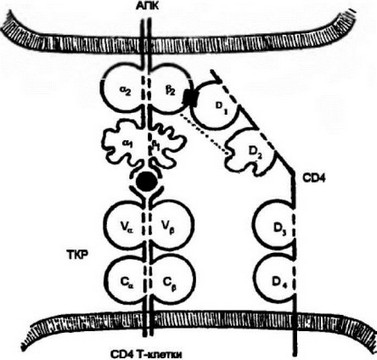
Рис. 3.17. Распознавание CD4 Т-клетками комплекса пе(гщд:мапекулы II класса.
После распознавания ТКР-комплекса пептидгмолекула II класса МНС в реакцию взаимодействия вступает ко рецептор CD4. Взаимодействие происходит между ^-доменом молекулы II класса и Б,-домена корецептора CD4. Корецептор имеет достаточно длинный цитоплазматический хвост, который передает сигнал о взаимодействии ТКР с антигенным комплексом внутрь клетки
ТКР антигенного комплекса происходит взаимодействие CD4 с молекулой II класса МНС. Реакция взаимодействия осуществляется между 02-доменом молекулы II класса МНС и первым доменом CD4. Предполагается также слабое включение во взаимодействие и второго домена — D2 .
Аналогичная картина наблюдается при распознавании антигенного комплекса цитотоксическими Т-лимфоцитами. Действующие участники взаимодействия: ТКР цитотоксических Т-лимфоцитов, комплекс пептида с молекулой I класса МНС и маркер цитотоксических Т-лимфоцитов — CD8 (рис. 3.18). CD8, хотя и выполняет сходную с CD4 функцию корецептора, структурно отличается от маркера Т-хелперов. Он представляет собой гетеродимер, каждая цепь которого включает один иммуноглобулинподоб- ный домен и достаточно длинный, связанный с мембраной участок цепи, который подвержен значительным конформационным изменениям. Также как и CD4, CD8 представлен на клеточной мембране независимо. Его функция корецептора реализуется в процессе антигенного распознавания. После взаимодействия ТКР
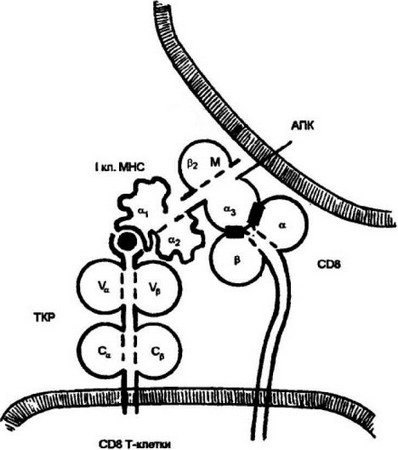
Рис. 3.18. Распознавание цитотоксическими Т-клетками (CD8 Т-клеткамн) комплексе пептид; молекулы 1 класса.
В процесс распознавания вступают ТКР и корецсптор CD8. CD8 взаимодействует своими а- и 0-доменами с аз-доменом молекулы I класса МНС. CD8, как и CD4, имеет относительно длинный цитоплазматический хвост, что позволяет ему передавать сигнал о взаимодействии внутрь клетки
с антигенным лигандом происходит контакт а- и р-доменов CD8 с а3-доменом молекулы I класса МНС. Образовавшийся молекулярный комплекс является условием передачи через корецептор CD8 сигнала внутрь клетки.
Внутриклеточные события, определяющие активацию Т-кле- ток, аналогичны тем, которые происходят в В-клетках после антигенной стимуляции. Образовавшийся агрегат из антигенпредстав- ляющих молекул МНС, Т-клеточного рецепторного комплекса, включающего CD3 молекулы, CD4 или CD8 молекул, провоцирует внутриклеточное взаимодействие различных тирозинкиназ с цитоплазматической частью полипептидов. Среди СОЗ-белков наибольшей связывающей активностью обладает CD3^, представленный в цитозоле не хвостовой, а головной частью. Активированные в результате взаимодействия киназы обеспечивают каскад реакций, следствием которых является индукция специфической транс- 108 крипции генов. Среди генов, вступивших в процесс транскрипции, особое место занимают те, которые кодируют синтез Т-за- висимых цитокинов, ИЛ-2 в частности. В конечном счете цепь событий от взаимодействия ТКР с антигенным комплексом и образования сложного молекулярного aiperara до внутриклеточных реакционных преобразований приводит к пролиферации и диффе- ренцировке Т-клеток до зрелых, функционально активных цитотоксических Т-лимфоцитов, или Т-хелперов.
Заключение
В- и Т-клетки обладают самостоятельными антигенраспознающими рецепторами, относящимися к одному и тому же су- персемейсгву иммуноглобулинов. Антигенраспознающие рецепторы В-клеток представляют собой мономерную форму IgM, модифицированную дополнительной последовательностью аминокислотных остатков в С-концевой части молекулы. Эта последовательность составляет трансмембранный и хвостовой участки тяжелой цепи. T-Клеточный антигенраспознающий рецептор (ТКР) состоит из двух полипептидных цепей, каждая из которых включает два домена: V и С. Механизм генетического контроля V-доменов, как slg, так и ТКР, в целом сходен и включает процесс случайной рекомбинации идентичных генных сегментов (V, D, J). Несмотря на то/ что функциональное предназначение антигенраспознаю- щих рецепторов двух типов клеток одно и то же — распознавание чужеродности, реализация такой функции В- и Т-клетками осуществляется разными способами. В то время как slg В-клеток распознает собственно антигенную детерминанту без каких-либо дополнительных условий, ТКР Т-клеток способен распознать только комплекс антигенной детерминанты с собственными молекулами I или II классов МНС.