
Распознавание мишеней фагоцитоза через различные мембранные рецепторы фагоцитов приводит к запуску процессов активации, отличающихся деталями, но приводящих к одинаковому результату — погружению и последующему разрушению частицы. Наиболее полно изучены активационные сигнальные процессы, запускаемые при адгезии опсонизированных клеток (особенно вариант передачи сигнала через FcyR). Этот и C3R-зависи- мый варианты активации фагоцитов схематично изображены на рис. 2.27.
На начальных стадиях фагоцитоза основные события происходят на обращенном к мишени участке поляризованной клетки, где должна сформироваться временная структура, называемая фагоцитарной чашей (phagocytic cap). На поверхности патогена обычно фиксировано несколько молекул антител. Они обусловливают объединение Fc-рецепторов в кластеры в результате перекрестного сшивания. Это приводит (за счет конформационных изменений) к активации прилежащих к цитоплазматической части рецепторов тирозинкиназ семейства Src. Эти киназы фосфорилируют цитоплазматические участки рецептора, в том числе остатки тирозина в мотиве ITAM, а также контактирующие с ними киназы семейства Syk. Фосфотирозины ITAM обеспечивают связывание этого мотива с S^-доменами
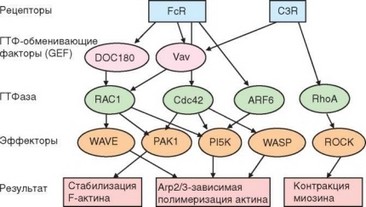
Рис. 2.27. Сигнальные пути, запускаемые связыванием рецепторов для опсонинов
Syk-киназ. Далее активационная волна передается на ряд ферментов, определяющих дальнейший ход событий.
На данном этапе наиболее важна активация двух ферментов: PI3K и PLC. PI3K катализирует превращение фосфатидилинозитол 4,5-дифосфата (PI4,5P2) в фосфатидилинозитол 3,4,5-трифосфат (PI3, 4,5P3), а PLC — расщепление PI4,5P2 с образованием диацилглицерола (DAG) и инозитолтри- фосфата. Диацилглицерол активирует протеинкиназу С. При этом уровень PI4, 5P2 не уменьшается благодаря его синтезу при участии фосфатидилино- зитол-4-фосфат-5-киназы. Выключение PI3K и PLC предотвращает развитие фагоцитоза.
Следующий этап активации направлен на образование продуктов, участвующих в полимеризации актина — процесс, на котором основан фагоцитоз. Для реализации этого этапа необходимо участие ГТФаз — Rac-1, Cdc42 (их активность преобладает при FcyR-зависимом фагоцитозе) и Rho (задействована преимущественно в комплемент-зависимом поглощении частицы). Нити актина окружают основание формирующейся фагоцитарной чаши. За их адгезию к мембране в этих участках отвечают белки семейства MARCKS (Myristoylated alanine-rich C kinase substrate), активируемые протеинкиназой С.
Активация ГТФаз происходит с участием белков обмена гуанидиновых нуклеотидов — GEF-белков (GEF — Guanine nucleotide exchanging factor). Участие в фагоцитозе наиболее четко показано для малого белка Vav, обладающего высоким сродством к PI3, 4, 5P3 и обеспечивающего активацию ГТФаз семейства Rho. В сборке F-актина основная роль принадлежит GEF-комплексу Arp1/2. Активацию этого комплекса осуществляет ГТФаза Cdc42 с участием белка WASP (Wiskott-Aldrich syndrome protein) — белок, связывающий нити актина с мембраной; дефект WASP приводит к развитию синдрома Вискотта—Олдрича — см. раздел 4.7.1.5). WASP выполняет роль связующего звена между прикрепленными к мембране фосфоинозитидами и GEF-комплексом Arp1/2. Полимеризацию актина контролируют и другие белки, однако их роль изучена мало.
Таким образом, полимеризация актина в сочетании с погружением частиц составляет основу формирования фагоцитарной чаши — главного результата рассмотренных процессов и исходной позиции следующего этапа фагоцитоза.
На начальных стадиях фагоцитоза основные события происходят на обращенном к мишени участке поляризованной клетки, где должна сформироваться временная структура, называемая фагоцитарной чашей (phagocytic cap). На поверхности патогена обычно фиксировано несколько молекул антител. Они обусловливают объединение Fc-рецепторов в кластеры в результате перекрестного сшивания. Это приводит (за счет конформационных изменений) к активации прилежащих к цитоплазматической части рецепторов тирозинкиназ семейства Src. Эти киназы фосфорилируют цитоплазматические участки рецептора, в том числе остатки тирозина в мотиве ITAM, а также контактирующие с ними киназы семейства Syk. Фосфотирозины ITAM обеспечивают связывание этого мотива с S^-доменами
Рис. 2.27. Сигнальные пути, запускаемые связыванием рецепторов для опсонинов
Syk-киназ. Далее активационная волна передается на ряд ферментов, определяющих дальнейший ход событий.
На данном этапе наиболее важна активация двух ферментов: PI3K и PLC. PI3K катализирует превращение фосфатидилинозитол 4,5-дифосфата (PI4,5P2) в фосфатидилинозитол 3,4,5-трифосфат (PI3, 4,5P3), а PLC — расщепление PI4,5P2 с образованием диацилглицерола (DAG) и инозитолтри- фосфата. Диацилглицерол активирует протеинкиназу С. При этом уровень PI4, 5P2 не уменьшается благодаря его синтезу при участии фосфатидилино- зитол-4-фосфат-5-киназы. Выключение PI3K и PLC предотвращает развитие фагоцитоза.
Следующий этап активации направлен на образование продуктов, участвующих в полимеризации актина — процесс, на котором основан фагоцитоз. Для реализации этого этапа необходимо участие ГТФаз — Rac-1, Cdc42 (их активность преобладает при FcyR-зависимом фагоцитозе) и Rho (задействована преимущественно в комплемент-зависимом поглощении частицы). Нити актина окружают основание формирующейся фагоцитарной чаши. За их адгезию к мембране в этих участках отвечают белки семейства MARCKS (Myristoylated alanine-rich C kinase substrate), активируемые протеинкиназой С.
Активация ГТФаз происходит с участием белков обмена гуанидиновых нуклеотидов — GEF-белков (GEF — Guanine nucleotide exchanging factor). Участие в фагоцитозе наиболее четко показано для малого белка Vav, обладающего высоким сродством к PI3, 4, 5P3 и обеспечивающего активацию ГТФаз семейства Rho. В сборке F-актина основная роль принадлежит GEF-комплексу Arp1/2. Активацию этого комплекса осуществляет ГТФаза Cdc42 с участием белка WASP (Wiskott-Aldrich syndrome protein) — белок, связывающий нити актина с мембраной; дефект WASP приводит к развитию синдрома Вискотта—Олдрича — см. раздел 4.7.1.5). WASP выполняет роль связующего звена между прикрепленными к мембране фосфоинозитидами и GEF-комплексом Arp1/2. Полимеризацию актина контролируют и другие белки, однако их роль изучена мало.
Таким образом, полимеризация актина в сочетании с погружением частиц составляет основу формирования фагоцитарной чаши — главного результата рассмотренных процессов и исходной позиции следующего этапа фагоцитоза.