
Хемокины (от Chemotactic cytokines) были открыты в конце 80-х годов прошлого столетия. Они составляют обширную группу цитокинов, объединенную сходным строением и способностью распознавать родопсиноподобные рецепторы.
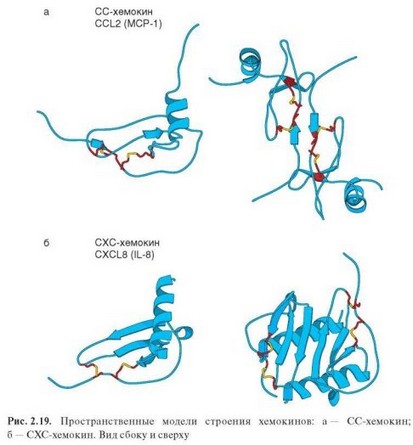
Хемокины — полипептиды молекулярной массой 8—12 кДа. Мономерные хемокины имеют сходную третичную структуру и состоят из индивидуального N-концевого участка, содержащего уникальную для данного хемо- кина последовательность, N-петли и сердцевинного домена, включающего р-складчатый участок (в а-хемокинах — 3 антипараллельные р-слоя), над которым расположена С-концевая а-спираль (рис. 2.19). Помимо сек- ретируемых, выделяют мембранные формы молекул хемокинов (например, фракталкин), выступающие также в роли молекул адгезии. Обычно хемо- кины присутствуют в биологических жидкостях в форме димеров, реже — тетрамеров. Четвертичная структура димеров существенно различается для хемокинов двух основных групп. Так, в а-хемокинах благодаря димеризации формируется большая плоскость, состоящая из p-слоев и «обрамленная» двумя а-спиралями (внешне напоминает поверхность для связывания пептидов в молекулах МНС). Хемокины способны формировать не только
гомо-, но и гетеродимеры. Функционально важное свойство хемокинов — их способность взаимодействовать с глюкозаминогликанами (гепарин, хондро- итинсульфат и др.) на поверхности клеток или в межклеточном матриксе. Иммобилизация хемокинов в тканях важна для создания их градиента, необходимого для направленного движения клеток. Индивидуальная N-концевая часть хемокинов может отщепляться металлопротеиназами. Видоизмененная таким образом молекула приобретает свойства ингибитора.
Выделяют 4 группы хемокинов. В основе разделения лежит взаимное расположение остатков цистеина (табл. 2.14). Всего в молекуле хемокинов имеется 4 остатка цистеина, образующих 2 дисульфидные связи. Два остатка цистеина, участвующие в формировании этих связей, расположены вблизи N-конца молекулы. В хемокинах одной группы, обозначаемой как а- или СХС-хемокины, упомянутые остатки цистеина (С) разделены любым аминокислотным остатком (Х). В хемокинах другой большой группы, обозначаемой как в- или СС-хемокины, остатки цистеина следуют друг за другом. Существует еще две минорные группы: у- или С-хемокины и 5- или СХ3С-хемокины. С-хемокины имеют всего 2 остатка цистеина, один из которых расположен в близи N-конца. В единственном представителе СХ3С-хемокинов (фракталкин) N-концевые остатки цистеина разделены 3 любыми остатками. Всего к настоящему времени описано 16 хемокинов группы СХС, 28 — группы СС, 2 — группы С и 1 — группы СХ3С. Разделение на группы отнюдь не формально. Внутри групп гомология первичной структуры значительно выше (достигает 70—90%), чем между группами (не превышает 40%). Рецепторы хемокинов в пределах группы сходны между собой и отличаются от рецепторов хемокинов другой группы. Хемокины разных групп различаются спектром мишеней: СС-хемокины привлекают моноциты и лимфоциты, но не нейтрофилы, а СХС-хемокины — нейтрофилы, реже лимфоциты, но не моноциты. Наконец, гены хемокинов у человека расположены в разных хромосомах: гены СХС-хемокинов — в 4q, CX — в 1q, СХ3С — в 16q, гены большинства CC-хемокинов — в 17q.
Среди СХС-хемокинов дополнительно выделяют 2 подгруппы, в зависимости от наличия в их молекуле перед первым остатком цистеина трипеп- тидной последовательности Glu—Leu—Arg (в однобуквенном коде — ELR). Эта последовательность характерна для восьми СХС-хемокинов (СХ^1— CXCL8) и отсутствует у остальных (СХСL9—СХCL18). Наличие последовательности ELR обусловливает способность а-хемокинов стимулировать ангиогенез (новообразование сосудов), воздействуя на эндотелиальные клетки.
В табл. 2.14 охарактеризованы представители основных групп хемокинов, а в табл. 2.15 — их рецепторы. Номенклатура хемокинов вначале создавалась стихийно, что вызывало большие трудности в связи с наличием у многих хемокинов синонимов. Именно поэтому недавно была введена единая классификация, основанная на обозначении групп (СС, СХС и т.д.), буквы L (лиганд, Ligand) и номера. Классификация хемокиновых рецепторов с самого начала была упорядочена; она включает обозначение группы, букву R (рецептор, Receptor) и номер. Выделяют 10 рецепторов СС-хемокинов, 7 рецепторов СХС-хемокинов и по 1 рецептору для хемокинов двух других групп.
Таблица 2.14. Характеристика хемокинов
|
Название |
Синонимы |
Молекулярная масса |
Рецепторы |
Клетки- мишени |
Функция |
|
СХС-хемокины |
|||||
|
CXCL1 |
GROa; MIP-2a |
7,9 |
CXCR2; CXCR1 |
Н |
Воспаление |
|
CXCL2 |
GROP; MIP-2P |
7,9 |
CXCR2 |
Н |
Воспаление |
|
CXCL3 |
GROy |
7,9 |
CXCR2 |
Н |
Воспаление |
|
CXCL4 |
PF-4 |
7,8 |
Нет данных |
ф |
? |
|
CXCL5 |
ENA-78 |
8,4 |
CXCR2 |
Н |
Воспаление |
|
CXCL6 |
GCP2 |
8,3 |
CXCR1; CXCR2 |
Н |
Воспаление |
|
CXCL7 |
NAP-2 |
7,6 |
CXCR1; CXCR2 |
Н |
Воспаление |
|
CXCL8 |
IL-8 |
8,4—8,9 |
CXCR1, CXCR2 |
Н |
Воспаление |
|
CXCL9 |
MIG |
11,7 |
CXCR3 |
Т-акт (Th1), НК, М, ЭК |
Гомеостаз, ангиогенез |
|
CXCL10 |
IP-10 |
8,6 |
CXCR3 |
Т-акт (Th1), НК, М, ЭК |
Гомеостаз, ангиогенез |
|
CXCL11 |
I-TAC |
8,3 |
CXCR3 |
Т-акт (Th1), НК, М, ЭК |
Гомеостаз, ангиогенез |
|
CXCL12 |
SDF-1 |
8,0;8,5 |
CXCR4 |
ЭК, все лейкоциты |
Гомеостаз, ангиогенез |
|
CXCL13 |
BCA, BLC |
10,3 |
CXCR5 |
В, ЭК |
Гомеостаз, ангиогенез |
|
CXCL14 |
BRAK |
9,4 |
Нет данных |
М, ЭК |
Воспаление, ангиогенез |
|
CXCL15* |
Lungkine |
Нет данных |
Нет данных |
Н, ЭК |
Воспаление, ангиогенез |
|
CXCL16 |
SIC |
10,2 |
CXCR6 |
Т-акт, ЭК |
Гомеостаз, ангиогенез |
|
СС-хемокины |
|||||
|
CCL1 |
I-309 |
8,5 |
CCR8 |
ДК, Т-акт (Th2) |
Воспаление |
|
CCL2 |
MCP-1 |
8,7 |
CCR2 |
В, М, Т-акт (Th1), НК, ДК |
Воспаление |
|
CCL3 |
MIP-1a |
7,7 |
CCR1, CCR2 |
Эо, Т-акт (Th1), М, НК, ДК |
Воспаление |
|
CCL4 |
MIP-1P |
7,8 |
CCR5 |
М, Т-акт (Th1), НК, ДК |
Воспаление |
|
CCL5 |
RANTES |
7,9 |
CCR1, CCR3, CCR5 |
Эо, В, Т-акт (Th1), М, НК, ДК |
Воспаление |
|
CCL6* |
C-10 |
|
CCR1, CCR2, CCR3 |
М, Т-акт (Th1) |
Воспаление |
|
CCL7 |
MCP-3 |
9,0 |
CCR1, CCR2, CCR3 |
Эо, В, Т-акт (Th1), М, НК, ДК |
Воспаление |
Окончание табл. 2.14
|
Название |
Синонимы |
Молекулярная масса |
Рецепторы |
Клетки- мишени |
Функция |
|
CCL8 |
MCP-2 |
8,9 |
CCR1, CCR3, CCR5 |
В, М, Т-акт (Th1), НК, ДК |
Воспаление |
|
CCL9/10[*] |
MIP-1y |
|
CCR1 |
Н, Т-акт |
Воспаление |
|
CCL11 |
Eotaxin |
8,4 |
CCR3 |
Эо, В, Т-акт (Th2), ДК |
Воспаление |
|
CCL12* |
MCP-5 |
|
CCR2 |
В, М, Т-акт, НК, ДК |
Воспаление |
|
CCL13 |
MCP-4 |
8,6 |
CCR1, CCR2, CCR3 |
Эо, В, Т-акт (Th1), М, НК, ДК |
Воспаление |
|
CCL14 |
HCC-1 |
8,7 |
CCR1 |
Эо, М |
Воспаление |
|
CCL15 |
HCC-2, MIP-1S |
9,2 |
CCR1, CCR3 |
Н, М, Эо, |
Воспаление |
|
CCL16 |
HCC-4, LEC |
11,2 |
CCR1 |
М, Т-акт, |
Воспаление |
|
CCL17 |
TARC |
8,1 |
CCR4 |
Т-акт (Th2) |
Воспаление, гомеостаз |
|
CCL18 |
PARC, MIP-4 |
7,9 |
CCR3 |
Т, ДК |
Гомеостаз |
|
CCL19 |
ELC |
8,8 |
CCR7 |
Т, ДК |
Гомеостаз |
|
CCL20 |
LARC |
8,0 |
CCR6 |
Т, В, ДК |
Гомеостаз |
|
CCL21 |
SLC |
12,3 |
CCR7 |
Т, ДК |
Гомеостаз |
|
CCL22 |
MDC |
8,1 |
CCR4 |
Т-акт (Th2), ДК |
Гомеостаз |
|
CCL23 |
MIP-3 |
11,4 |
CCR1 |
Н, М, Т |
Воспаление |
|
CCL24 |
Eotaxin-2 |
10,5 |
CCR3 |
Эо, В, Т-акт (Th2), ДК |
Воспаление |
|
CCL25 |
TECK |
14,2 |
CCR9 |
Т, тимоциты |
Гомеостаз |
|
CCL26 |
Eotaxin-3 |
8,4 |
CCR3 |
Эо, В, Т-акт (Th2), ДК |
Воспаление |
|
CCL27 |
CTACK |
10,2 |
CCR3, CCR2, CCR10 |
Т-акт |
Гомеостаз |
|
CCL28 |
MEC |
12,4 |
CCR10 |
Т-акт |
Гомеостаз |
|
C-хемокины |
|||||
|
XCL1 |
Lymphotactin |
10,3 |
XCR1 |
T |
Гомеостаз |
|
XCL2 |
SCM-1P |
10,3 |
XCR2 |
T |
Гомеостаз |
|
CXjC-хемокины |
|||||
|
СХ3СК1 |
Fractalkine |
8,6 |
CX3CR1 |
Т-акт, НК, М |
Гомеостаз |
Таблица 2.15. Рецепторы хемокинов
|
Рецептор |
Лиганды |
Распределение |
|
CXCR1 |
CXCL6, CXCL7, CXCL8 |
Нейтрофилы |
|
CXCR2 |
CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7, CXCL8 |
Нейтрофилы |
|
CXCR3 |
CXCL9, CXCL10, CXCL11 |
Нейтрофилы, ТЫ-клетки |
|
CXCR4 |
CXCL12 |
Все лейкоциты |
|
CXCR5 |
CXCL13 |
В-лимфоциты |
|
CXCR6 |
CXCL16 |
Активированные Т-клетки |
|
CXCR7 |
CXCL12 |
Все лейкоциты |
|
CCR1 |
CCL3, CCL5, CCL7, CCL13, CCL14, CCL15, CCL16, CCL23 |
Моноциты, эозинофилы, дендритные клетки, NK-клетки, активированные Т-клетки |
|
CCR2 |
CCL2, CCL7, CCL8, CCL13, CCL16 |
Моноциты, эозинофилы, дендритные клетки, NK-клетки, активированные Т-клетки (Th1) |
|
CCR3 |
CCL5, CCL7, CCL8, CCL11, CCL13, CCL15, CCL16, CCL24, CCL26, CCL28 |
Моноциты, эозинофилы, дендритные клетки, NK-клетки, активированные Т-клетки (Th2) |
|
CCR4 |
CCL17, CCL22 |
^2-клетки |
|
CCR5 |
CCL3, CCL4, CCL5, CCL8, CCL11, CCL14, CCL16 |
Моноциты, эозинофилы, дендритные клетки, NK-клетки, активированные Т-клетки (Th1) |
|
CCR6 |
CCL20 |
Дендритные клетки, Т- и В-лимфо- циты |
|
CCR7 |
CCL19, CCL21 |
Дендритные клетки (зрелые), наивные Т-клетки |
|
CCR8 |
CCL1 |
Дендритные клетки, ^2-клетки |
|
CCR9 |
CCL25 |
Тимоциты, Т-клетки |
|
CCR10 |
CCL27, CCL28 |
Активированные Т-клетки |
|
XCR1 |
XCL1, XCL2 |
Т-клетки |
|
CX3CR1 |
CX3CL1 |
Моноциты, активированные Т-клет- ки, NK-клетки |
По функциональной роли выделяют:
- гомеостатические хемокины;
- провоспалительные хемокины.
Гомеостатические хемокины отвечают за распределение клеток (прежде всего лимфоцитов) по лимфоидным органам. К этим цитокинам относят CXCL12, CXCL13, CCL17, CCL19, CCL20, CCL21, CCL22, CCL25, CCL27, CCL28, а также CX3CL, XCL1 и XCL2. Остальные хемокины (подавляющее большинство) относят к группе провоспалительных, поскольку они отвечают за активацию клеток и привлечение их в очаг воспаления. Хемокины этих групп различаются условиями синтеза и секреции: гомеостатические цитокины секретируются постоянно, обычно стромальными и эндотели
альными, иногда дендритными клетками, реже — самими лимфоцитами. Хемокины определяют миграцию клеток иммунной системы в процессе их созревания. Так, хемокины обусловливают «заселение» тимуса лимфоидными клетками-предшественниками, направление миграции тимоцитов в процессе созревания внутри тимуса и последующую эмиграцию Т-кле- ток в периферические отделы иммунной системы. Аналогично хемокины определяют направление миграции дендритных клеток на разных этапах их созревания: сначала — движение клеток из кровотока в барьерные ткани, а затем перемещение в Т-зоны региональных лимфатических узлов. В лимфоидных органах разные участки стромы секретируют различные гомеостатические хемокины, что определяет их заселение разными типами лимфоцитов (Т-зоны — Т-клетками, а В-зоны — В-клетками). Это обеспечивает стабильность состава лимфоидных органов при непрерывной рециркуляции лимфоцитов.
Хемокиновые рецепторы, как и рецепторы других «профессиональных» хемотаксических агентов, относят к семейству родопсиноподобных рецепторов. Они 7-кратно пронизывает мембрану а-спирализованными участками (7-трансмембранные рецепторы) (см. рис. 2.20). N-конец молекулы рецептора и 3 петли обращены наружу и участвуют (особенно N-конец) в распознавании и связывании хемокина-лиганда. С-конец и 3 петли обращены внутрь клетки. С внутриклеточными петлями рецептора связаны G-белки (белки, связывающие гуаниннуклеотиды), что определяет обозначение этих рецепторов как GPCR (Gprotein-coupled receptor). Роль G-белков в передаче хемо- киновых сигналов в клетку будет рассмотрена далее. Хемокиновые рецепторы обладают высоким сродством к лигандам (порядка 10-9 М). Сродство основано на взаимодействии домена сердцевины хемокина с N-концевой
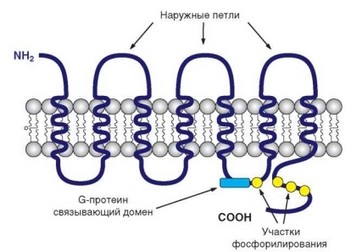
Рис. 2.20. Структура рецептора для хемокинов, его положение в клеточной мембране и связь с G-белком
частью рецептора. Пептиды, воспроизводящие фрагменты этих участков хемокина и рецептора, используют как инструменты для модификации эффектов хемокинов. Специфичность хемокиновых рецепторов характеризуется вырожденностью: с одним и тем же рецептором взаимодействует до 10 хемокинов (например, в случае CCR3). Только единичные рецепторы имеют один лиганд (см. табл. 2.15). С другой стороны, системе хемокинов свойственна избыточность: один и тот же хемокин может взаимодействовать с разными рецепторами, как, например, CXCL8/IL-8, распознаваемый рецепторами CXCR1 и CXCR2. Как правило, гомеостатические и провоспалительные хемокины взаимодействуют с разными рецепторами.
При взаимодействии хемокинов с их рецепторами на поверхности клетки происходит димеризация рецепторов. Вслед за этим запускаются события, вовлекающие G-белки, связанные с гуаниннуклеотидами. Функция G-белков состоит в регуляции внутриклеточных сигнальных процессов, осуществляемой за счет обратимых переходов ГТФ^ГДФ. G-белки чувствительны к токсину столбняка, инактивирующему передачу сигнала от этих рецепторов внутрь клетки, обусловленную обменом ГТФ^ГДФ и молекулярными превращениями белка G. G-белки — тримерные молекулы, содержащие цепи Ga, Gp и Gy. Ga-цепь связана с ГДФ. Конформационные изменения, происходящие в рецепторе при связывании хемокина, и его фосфорилирование под влиянием тирозинкиназ обусловливают замещение ГДФ на ГТФ, что вызывает выход Ga-цепи из состава комплекса и переход ее в цитозоль. Gp- и Gy-цепи остаются в составе димера, связанного с мембраной. Разделившиеся компоненты G-белка действуют затем самостоятельно (рис. 2.21). Их эффекты состоят в активации ряда протеинкиназ (тирозинкиназы семейства Src, серинтреониновой протеинкиназы С, липидной фосфатидилинозитол 3-киназы), а также ГТФ-свзывающих белков и ГТФаз.
Достаточно подробно изучена роль димера Ру. Он активирует 2 фермента — фосфолипазу С (PLC) (изоформа Р) и фосфатидилинозитол-3-киназу (PI3K, изоформа у). Эти ферменты участвуют в функционировании различных сигнальных путей. Важнейший продукт, формирующийся из мембранных фосфоинозитидов под влиянием PLC, — инозитол 3,4,5-трифосфат. Действуя на рецепторы на поверхности мембраны эндоплазмтического ретикулума, он вызывает раскрытие кальциевых каналов и выход в цитозоль депонированного Са2+. Другой продукт превращения фосфоинозитидов — диацил- глицерол — активирует некоторые изоформы серинтреониновой протеинкиназы С, участвующей во многих внутриклеточных процессах. Это звено важно для обеспечения активации клетки, но непосредственно не связано с основным результатом действия хемотаксических факторов — влиянием на цитоскелет. Оно будет рассмотрено более подробно в разделе 3.5.2.1.
Активация цитоскелета происходит с вовлечением сигнальных путей, зависимых и не зависимых от PI-3^ Фосфатидилинозитол-3-фосфат активирует протинкиназу В (Akt), а также низкомолекулярные гуанининнукле- оидсвязывающие белки семейства Rh0, (Rho, Rap1 и Cdc42); их называют факторами обмена гуаниннуклеотидов — GEF (Guanine-nucleotide exchange factors). Обеспечивая превращение ГДФ в ГТФ эти белки активируют малую ГТФазу Rac — незаменимый фактор клеточной миграции. Rac реализует свою активность через взаимодействие с факторами, контролирующими
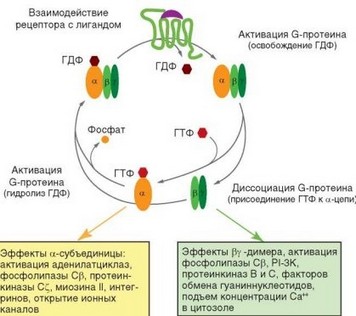
Рис. 2.21. Активация и интактивация G-белков, связанных с рецепторами, и вызываемые ими биологические эффекты
актин (WASP, WIP, Arp1/2, миозин II). В результате происходит полимеризация мономерного G-актина с его превращением в F-актин (филамен- тозный) (рис. 2.22). Актин составляет основу микрофиламентов и служит главным контрактильным белком клетки, от которого зависит ее движение, формирование контактов и фагоцитоз. Направленность движения клетки определяется активностью GEF-белка Cdc42, обеспечивающего формирование F-актина и удаление некоторых ингибиторов из лидирующего края клетки. Для проявления Cdc42 своей активности необходим белок РАС-1. В результате локального образования F-актина формируется ламеллоподия, в которую перемещается значительная часть актина и действующих факторов локомоции. Поляризация клетки и активация интегринов (обеспечивает усиление адгезии клеток к субстрату и объектам фагоцитоза) происходит при участии еще одного из упомянутых GEF-белков — Rар1.
Роль комплекса а-цепи G-белка с ГТФ изучена менее подробно; ее можно расценивать как вспомогательную. Комплекс участвует в подготовке клетки к миграции. При этом происходит активация Rho-GEF, участвующего в активации изоформы Z протеинкиназы С (контролирует состояние интегринов), а также в активации контрактильного белка миозина II, что необходимо для реализации сократительной активности микрофиламентов. Помимо роли в контракции цитосеклета, комплекс а-цепи и ГТФ участвует

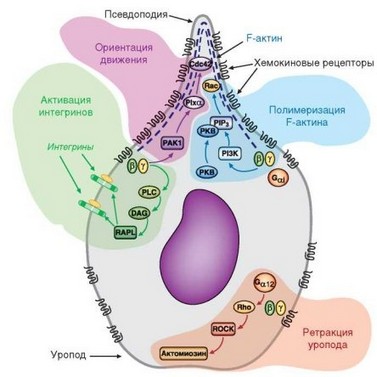
в активации интегринов. Поскольку Ga-цепь обладает слабой ГТФазной активностью, ГТФ в составе комплекса превращается в ГДФ, и Ga-цепь воссоединяется с димером GPy. Образующийся тример восстанавливает свою связь с хемокиновым рецептором и передача сигнала прекращается.
Таким образом, связывание хемотаксических факторов с рецепторами сопряжено с активацией интегринов, усилением адгезии, поляризацией клеток, образованием полимерной формы актива (F-актина), формированием двигательного псевдоподия (ламеллоподия) и, наконец, контракцией актомиозина, обеспечивающей движение клетки.