
В обеспечении киллинга фагоцитированных микроорганизмов наиболее важна роль производных кислорода. Главное событие в образовании кислородзависимых бактерицидных факторов — кислородный взрыв — быстрое (реализуемое за секунды) и высокопродуктивное осуществление цепи реакций, приводящих к образованию активных форм кислорода. Активные формы кислорода включают высокореактивные свободные радикалы, ионы кислорода и кислородсодержащих химических групп. Образование активных форм кислорода катализируется ферментом NADPH-оксидазой (NADPH — восстановленная форма никотинамиддинуклеотидфосфата), называемой также оксидазой фагоцитов (P^x).
Сборка NADPH-оксидазы
Кислородный взрыв реализуется в мембране фаголизосом. Сборка NADPH-оксидазы — исходное событие в запуске кислородного взрыва и генерации активных форм кислорода. NADPH-оксидаза активируется под влиянием сигналов, возникающих при связывании лигандов с Fc-рецепторами, рецепторами-«мусорщиками», родопсиноподобными рецепторами. Сначала этот процесс происходит в участке клеточной мембраны, примыкающем к зоне действия хемотаксического агента или к зоне контакта с фагоцитируемым объектом. При формировании фагосомы сборка NADPH- оксидазы перемещается внутрь клетки.
Неактивная NADPH-оксидаза локализуется в мембране секреторных везикул и специфических гранул. Она представляет собой флавоцитохром b558, состоящий из субъединиц a (p22phox) и в (gp91phox), а также белка Rap^. При активации к NADPH-оксидазе подсоединяется 4 цитозольных белка: p47phox, p67phox, p40phox и ГТФаза Rac-2. При фагоцитозе гранулы и везикулы, содержащие NADPH-оксидазу, сливаются с фагосомой. Активация фермента происходит уже в составе мембраны фаголизосомы. В привлечении цитозольных компонентов важная роль принадлежит мембранным фосфоинозитидам, обеспечивающим связывание цитозольных компонентов Phox — фосфотидилино- зитол-3,4-дифосфата (PI-3,4P2) и фосфотидилинозитол-3-монофосфата (PI-3P), а также фосфотидилинозитолкиназ, катализирующих их образование.
Под влиянием активационных сигналов, генерируемых при связывании родопсиноподобных рецепторов, происходят фосфорилирование p47phox и активация двух его доменов: PX и SH3. При участии PX-домена p47phox прикрепляется к PI(34)P2 мембраны фаголизосомы, а с помощью SH3-домена — к С-кон- цевой цитозольной части мембранного компонента цитохрома b558 — p22phox. Переместившись в мембрану, p47phox способствует внедрению в нее белка p67phox, происходящему за счет взаимодействия С-концевого домена p47phox и С-кон- цевого SH3-домена p67phox. Адапторный белок p40phox связан с p67phox исходно, а через PX-домен он присоединяется к PI-3P мембраны. Одновременно ГТФаза Rac-2 освобождается от ингибитора, блокирующего ее при покоящемся состоянии клетки (отделение от ингибитора обусловлено фосфорилированием Rac-2), и внедряется в мембрану фагосомы, где взаимодействует с молекулой p67phox. Таким образом, осуществляется полная сборка NADPH-оксидазы (рис. 2.28).
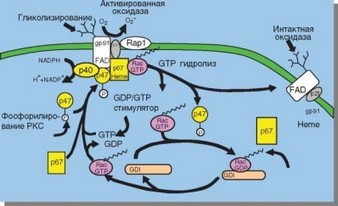
Рис. 2.28. Схема активации NADPH-оксидазы
Образование активных форм кислорода
Конформационные изменения, происходящие при сборке NADPH-окси- дазы, приводят к тому, что ее основной компонент gp91phox приобретает способность взаимодействовать с окисленной формой кофактора, образующегося при гликолизе, — NADPH. Это взаимодействие происходит при участии простетической группы FAD (флавин адениндинуклеотид) и двух молекул гема. FAD получает электрон (е-) от NADPH и передает его «наружной» молекуле гема, обращенной к цитозолю, от которой он переходит к «внутренней» молекуле гема, обращенной к содержимому фагосомы. Внутренняя молекула гема передает электрон молекуле кислорода, что приводит к образованию супероксида, объединяющего в себе свойства аниона и радикала, и потому называемого супероксидрадикалом, или супероксиданионом (*О21). Супероксидрадикал — короткоживущий родоначальник активных форм кислорода (рис. 2.29). В начальную фазу фагоцитоза супероксиданион, образующийся на участке клеточной мембраны, находится во внутриклеточном пространстве; после формирования фагосомы и фаголизосомы он поступает внутрь этих гранул.
На следующем этапе реализуется цепь реакций, приводящих к образованию радикалов, ионов кислорода и содержащих их молекул, обладающих более высокой бактерицидной активностью чем супероксианион, — активных форм кислорода. Под действием фермента супероксиддисмутазы из двух молекул супероксидного аниона образуется перекись водорода. В присутствии ионов Fe2+ супероксид взаимодействует с перекисью водорода
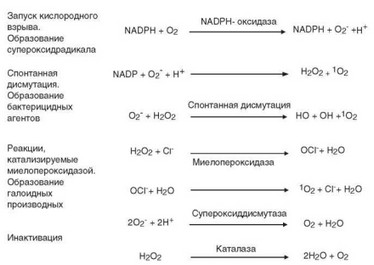
Рис. 2.29. Кислородзависимые процессы в фагоцитах, приводящие к образованию бактерицидных веществ — активных форм кислорода и галоидсодержащих метаболитов. Слева — основные стадии процесса; справа — химические реакции и их продукты
с образованием гидроксил-радикала (*ОН) (см. рис. 2.29) — сильного окислителя. Перекись водорода и особенно гидрокисл-радикал обладают очень сильной бактерицидной активностью. При их совместном действии происходит перекисное окисление липидов, разрыв пептидных связей, окисление сульфгидрильных групп и другие глубокие химические изменения макромолекул в клеточных стенках патогенов, приводящие к их гибели. При мутациях генов, кодирующих субъединицы NADPH-оксидазы, нарушается активность этого фермента и, как следствие, развивается хроническая гранулематозная болезнь (см. раздел 4.7.1.4).
Кислородзависимые факторы бактерицидности, индуцируемые миелопероксидазой
Миелопероксидаза — маркерный фермент азурофильных гранул нейтрофилов. Она составляет 1—5% общего белка этих клеток. Зрелая молекула миелопероксидазы — гетеродимер, образованный тяжелой а- и легкой p-цепями. С а-цепью связана железосодержащая группа — гем. Миелопероксидазная микробицидная система включает, помимо собственно миелопероксидазы, перекись водорода и кофакторы, в том числе ионы галогенов (Cl-, I-, Br-).
Миелопероксидаза катализирует в фаголизосомах окислительные реакции. Превращение йодида (I-) в молекулярный йод (при участии миелопе- роксидазы и перекиси водорода) обеспечивает его связывание с сульфгид- рильными группами белков, приводящее к нарушению жизнеспособности микроорганизмов. При катализируемом миелопероксидазой взаимодействии ионов Cl- с перекисью водорода образуется сильный микробицидный агент — хлорноватистая (гипохлорная) кислота HOCl. При ее взаимодействии с аминокислотами образуются хлорамины, обладающие бактерицидным действием. При окислении хлорноватистой кислоты супероксидом образуется гидроксильный радикал *ОН, а при ее оксилении перекисью водорода — синглетный кислород ‘О2 (см. рис. 2.29) Эти метаболиты обладают сильной микробицидной активностью. Синглетный кислород особенно активно взаимодействует с полиненасыщенными жирными кислотами, вызывая их перекисное окисление, нарушающее целостность мембраны бактерий. Синглетный кислород участвует в образовании еще одного микробицид- ного вещества — озона (О3).
Нейтрофилы — наиболее эффективные продуценты активных форм кислорода. К этим агентам чувствительны разные типы микроорганизмов, в первую очередь — внеклеточные патогены.