
Некоторые из молекул, связанные с распознаванием объектов фагоцитоза, упомянуты выше. Рассмотрим 2 типа рецепторов, играющих основную роль в распознавании опсонинов, фиксированных на поверхности фагоцитируемых клеток: рецепторы для Fc-части иммуноглобулинов/антител (FcR) и рецепторы для комплемента (CR).
Fc-рецепторы
Распознавание патогенов и других клеток, опсонизированных антителами класса IgG, осуществляется с помощью Fc-рецепторов (FcyR), экспрессированных на поверхности фагоцитов. Эти рецепторы распознают участки хвостовой части (Fc, см. раздел 3.1.1.1) молекул иммуноглобулинов IgG-класса (в наибольшей степени субклассов IgG1 и IgG3). Эти участки расположены в ^2- и СН3-доменах у-цепей иммуноглобулинов. В молекулах свободных антител они скрыты и открываются только при связывании с антигеном, сопровождающимся изменением конфигурации молекулы IgG. Кроме Fcy-рецепторов, известны Fc-рецепторы, распознающие связанные антитела классов IgA и IgE (FcaR и FceR). Функциональная роль Fc-рецепторов значительно шире их участия в распознавании объектов фагоцитоза, поэтому они будут многократно упоминаться в дальнейшем. Основные характеристики Fc-рецепторов представлены в табл. 2.18 и на рис. 2.26.
У человека выделяют 3 типа Fcy-рецепторов — FcyRI (CD64), FcyRII (CD32) и Fc yRIII (CD16); 2 последних типа существуют в нескольких вариантах, различающихся устройством С-концевой части. Перечисленные рецепторы отличаются друг от друга сродством к Fc-части молекулы IgG. В порядке убывания аффинности они образуют ряд: FcyRI gt; FcyRII gt; FcyRIII.
| Традиционное обозначение | CD-нумера
ция |
Молекулярная масса, кДа | Экспрессия на клетках | Сродство к изотипам иммуноглобулинов | Аффинитет (Kd), M |
| FcyRI | CD64 | 72-75 | Моноциты/макрофаги, активированные нейтрофилы, фолликулярные дендритные клетки | IgGl=IgG3 gt;IgG2 gt;IgG4 | IQ-S-IQ-9* |
| FcyRII | CD32 | 40-50 | Моноциты/макрофаги, нейтрофилы, В-клетки, эозинофилы, тромбоциты, дендритные клетки, активированные Т-клетки | IgGl=IgG3 gt;IgG3 gt;IgG4 | 1СГ7 |
| FcyRIII | CD16 | 50-65 | Естественные киллеры, моноциты/макрофаги, активированные нейтрофилы, активированные эозинофилы, активированные Т-клетки | IgGl, IgG3 | 1СГ6 |
| FcaR | CD89 | 55-75 | Т1ейтрофилы, активированные моноциты/макрофаги, активированные эозинофилы, Т-клет- ки, В-клетки | IgA | Нет данных |
| FceRI | a-29,6; P — 26,5; У-9,7 | Тучные клетки, В-клетки, дендритные клетки, эозинофилы | IgE | 1(Г9-1(Г10* | |
| FceRII | CD23 | 45-50 | В клетки, активированные моноциты/макрофаги, эозинофилы, активированные Т-клет- ки, фолликулярные дендритные клетки | IgE | КГ7 |
Таблица 2.18. Характеристика Fc-рецепторов
* — способны связывать нативные (не в составе комплекса с антигеном) молекулы антител.
2.3. Клеточные механизмы врожденного иммунитета 127
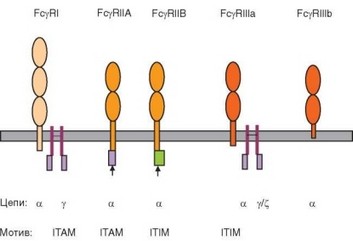
Рис. 2.26. Строение мембранных Fc-рецепторов. Особо акцентировано наличие в цитоплазматической части рецепторов мотивов, передающих активационные (ITAM) или ингибирующие (ITIM) сигналы. Линии, соединяющие изображения цепей, обозначают дисульфидные связи
Наиболее высокоаффинный рецептор FcyRI состоит из двух полипептидных цепей. Цепь, связывающаяcя c Fc-участком у-цепи IgG (а-цепь), имеет 3 внеклеточных домена, относящихся к суперсемейству иммуноглобулинов. Ее цитоплазматический участок лишен последовательностей, позволяющих передавать сигнал внутрь клетки. Такая активационная сигнальная последовательность (участок ITAM — Immunoreceptor tyrosine-based activation motif) есть в цитоплазматической части другой полипептидной цепи — у-цепи, осуществляющей в связи с этим сигнальную функцию рецептора. FcyRI экспрессирован на покоящихся клетках только одного типа — макрофагах; нейтрофилы и эозинофилы начинают экспрессировать его после активации. Это единственный тип Fcy-рецепторов, связывающий свободные антитела. Фиксация антител на FcyRI обусловливает формирование «армированных» макрофагов, несущих на поверхности антитела с активными центрами, направленными наружу.
Рецепторы FcyRII представлены более широко: они свойственны практически всем клеткам врожденного иммунитета, а также В-лимфоцитам. FcyRII тоже имеют одну а-цепь, содержащую 2 внеклеточных иммуно- глобулинподобных домена. Внутриклеточный участок а-цепи FcyRII по протяженности превосходит аналогичные участки других Fc-рецепторов. Известно 2 варианта этих рецепторов — FcyRIIA и FcyRI№, различающися особенностями строения их цитоплазматической части. FcyRIIA содержит в ней активационный мотив ITAM, а FcyRI№ — ингибирующий мотив
ITIM (Immunoreceptor tyrosine-based inhibition motif). Фосфорилирование остатков тирозина в ITAM делает возможным взаимодействие его с тирозинкиназами семейства Syk, что служит основой для передачи активационного сигнала (см. раздел 3.5.2.1). Фосфорилирование тирозина в ITIM обеспечивает его взаимодействие с фосфатазами SHP1, SHP2 и SHIP, ослабляющими активационный сигнал и оказывающими ингибирующее действие (см. раздел 3.6.6.3). Рецепторы FcyRIIA присутствуют на поверхности фагоцитов различной природы и играют важную роль в распознавании опсонизирующих антител. Рецепторы FcyRIIB экспрессированы преимущественно на В-лимфоцитах и участвуют в регуляции их активности.
Низкоаффинные рецепторы FcyRIII имеют сложную структуру. Помимо основной a-цепи, имеющей 2 иммуноглобулинподобных внеклеточных домена, они содержат 2 дополнительные цепи, идентичные у-цепям рецепторов FcyRI или FceRI. Эти цепи несут участок ITAM. По особенностям структуры С-концевой части a-цепи эти рецепторы также неоднородны. Их основной вариант — FcyRIIA — содержит a-цепь, имеющую полноценную трансмембранную и внутриклеточную части, причем последняя взаимодействует с у-цепью, что позволяет рецептору передавать сигнал в клетку. Такой вариант рецептора характерен для естественных киллерных клеток, а также моноцитов, макрофагов и ряда других клеток. Альтернативный вариант — FcyRIIK, содержащий a-цепь, заякоренную в мембране через гликозилфосфатидилинозитол. Этот вариант рецептора экспрессируется на нейтрофилах.
СЗ-рецепторы
С3-рецепторы по своей структуре более гетерогенны, чем Fc-рецепторы. Известно 4 разновидности С3-рецепторов (табл. 2.19). CR1 (CD35) экспрессирован не только на классических фагоцитах (нейтрофилах, моноцитах), но и на ряде других клеток: В-лимфоцитах, эритроцитах, фолликулярных дендритных клетках. На последних CR1 участвует в связывании иммунных комплексов, т.е. выполняет важнейшую функцию, играющую ключевую роль в развитии гуморального иммунного ответа (см. раздел 3.5.2.2). Молекула CD35 связывает, помимо C3b, фрагмент C3d. Рецептор CR2 (CD21) имеет наиболее широкий спектр лигандов. Он связывает фрагменты С3, находящиеся на разных стадиях деградации — С3Ь, iC3b, С3d. Кроме того, CR2 служит рецептором вируса Эпштейна—Барр. Этот рецептор экспрессирован на В-лимфоцитах и фолликулярных дендритных клетках, но не на фагоцитах. Он не имеет отношения к фагоцитозу, но играет важную роль в активации В-клеток (см. раздел 3.6.2.1) и процессах гуморального ответа, происходящих в зародышевых центрах. Наконец CR3 и CR4 представляют уже охарактеризованные (см. раздел 2.3.1) р2-интегины — Мас-1 (CD11b/CD18) и p150,95 (CD11c/CD18). Они связывают инактивированный фрагмент C3b — iC3b (см. раздел 2.5.1). Все С3-рецепторы играют важную роль в контроле активации комплемента, ингибируя связывание С3 с поверхностью собственных клеток и ускоряя отделение компонентов комплемента от иммунных комплексов.
Таблица 2.19. Рецепторы для компонентов комплемента на клетках человека
|
Название |
Лиганды |
Молекулярная масса, кДа |
Экспрессирующие клетки |
Биологические функции |
|
CR1 (CD35) |
C3b, iC3b, C4b, C3d |
160-250 |
Эритроциты, моно- циты/макрофаги |
Опсонизация, расщепление С3Ь, клиренс иммунных комплексов |
|
CR2 (CD21) |
iC3b, C3dg, C3d |
140 |
В-клетки, Т-клетки, естественные киллеры |
Иммунорегуляция, связывание вируса Эпштейна-Барр |
|
CR3 (CD11b/ CD18) |
iC3b |
170/95 |
Моноциты/макрофа- ги, нейтрофилы |
Опсонизация, расщепление iC3b |
|
CR4 (CD11c/ CD18) |
C3b, iC3b, C3dg |
150/95 |
Моноциты/макрофа- ги, нейтрофилы |
Опсонизация |
|
C3aR |
C3a |
59 |
Тучные клетки, базофилы, моноциты/ макрофаги, нейтрофилы |
Освобождение медиаторов, хемотаксис |
|
C5aR |
C5a |
43 |
Нейтрофилы, моноциты, базофилы, эозинофилы |
Освобождение медиаторов, хемотаксис |
|
С5L2 |
C5a |
41 |
Нейтрофилы, незрелые дендритные клетки |
То же; выражено слабее |