
Лимфатические узлы — бобовидные образования величиной 0,2—1,0 см, образующие группы или цепочки. Они дренируют лимфу от различных регионов тела. У мышей выделяют следующие лимфатические узлы: нижнечелюстные, шейные, подмышечные, брыжеечные, поясничные, крестцовые, подвздошные, паховые, подколенные. В эмбриогенезе они формируются не одновременно. У мышей ранее других (на 9-10-е сутки эмбрионального развития) закладываются брыжеечные, крестцовые и шейные лимфатические узлы, а позже всех (на 15—17-е сутки) — подвздошные и подколенные узлы. Остальные лимфатические узлы формируются между 10 и 15-ми сутками.
Структура лимфатических узлов схематически изображена на рис. 3.68. Со стороны выпуклой части лимфоузла через трабекулы в орган входят лимфатические сосуды, приносящие лимфу из дренируемых участков тела. В своей вогнутой части узел имеет ворота, через которые входят афферентные артерии и выходят эфферентные лимфатические сосуды и вены. Лимфатический узел окружен соединительнотканной капсулой, от которой внутрь отходят тонкие перегородки — трабекулы. Ткань лимфатического узла состоит из коры и мозгового вещества. В наружной части коры есть округлые образования — лимфоидные фолликулы, служащие местом сосредоточения В-лимфоцитов. В покоящихся лимфоузлах они имеют равномерную структуру, и их обозначают как первичные фолликулы. При иммунном ответе в фолликул, помимо В-лимфоцитов, мигрируют Т-клетки, в нем формируются центры размножения — зародышевые центры. Такой фолликул называют вторичным. Ключевую роль в формировании и функционировании фолликулов и развивающихся в них зародышевых центров играют фолликулярные дендритные клетки, по происхождению не имеющие отношения к обычным дендритным клеткам и сходные
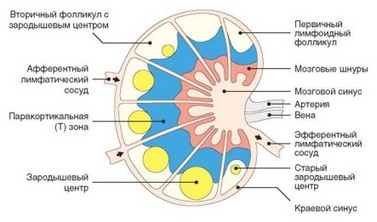
Рис. 3.68. Строение лимфатического узла и локализация в нем Т- и В-лимфоцитов
(по Janeway С.А. et al., 2005)
с ними только по морфологии. Эти клетки имеют местное мезенхимальное происхождение.
Формирование Т- и В-зон лимфатических узлов, как и других вторичных лимфоидных органов, а также мозгового слоя тимуса определяется взаимодействием лимфоидных клеток и предшественников стромальных клеток, осуществляемое в эмбриогенезе. Ключевая роль в нем принадлежит мембранным лимфотоксинам (LT) — цитокинам семейства TNF — и их рецепторам. Из костного мозга в зачаток лимфатических узлов и других лимфоидных органов мигрируют специальные лимфоидные клетки, обозначаемые как клетки-индукторы лимфоидной ткани — LTIC (Lymphoid tissue inducing cells). Их мембранный фенотип — CD4+ CD3- CD45+ а4Р7+ IL-7Ra+ LTaP+ CXCR5+. LTIC экспрессируют дифференцировочный фактор RORyt (Retinoid-acid-receptor-related orphan receptor — орфановый рецептор, родственный рецептору ретиноидной кислоты) и транскрипционный фактор Id2. Судя по мембранным молекулам, эти клетки имеют костномозговое лимфоидное происхождение, но не относятся к зрелым субпопуляциям лимфоцитов. LTIC содержат на своей поверхности мембранную форму LTaP — тример, включающий 2 трансмембранные молекулы LTp и 1 молекулу ЕГа, расположенную внеклеточно. Другая сигнальная молекула LTIC, также относящаяся к семейству TNF, — RANKL (Recerptor activator of NFKB-ligand). В образовании лимфоидных органов участвуют также клетки-инициаторы лимфоидной ткани — LTIN, которые имеют фенотип CD4- CD3- IL-7Ra- CD11b+ CD11c+, свидетельствующий об их родстве с дендритными клетками. С этими двумя типами клеток взаимодействуют предшественники стромальных клеток — клетки-организаторы лимфоидной ткани — Lto.
LTp и RANKL распознаются рецепторами семейства TNFR на стромальных клетках, в результате чего в них генерируется сигнал, вызывающий экспрессию генов IL-7, хемокинов (CXCL13, CCL19, CCL21) и фактора TRANCE (TNF-related activation-induced cytokine). Все эти факторы действуют на лимфоидные клетки; кроме того, они привлекают зрелые лимфоциты, обеспечивают их жизнеспособность и способствуют их кластеризации с образованием зачатка фолликула. Затем мигрирующие в зачаток В-, Т- и NK-лимфоциты разделяются пространственно. К этому моменту в формирующемся органе появляются дендритные клетки, играющие роль организующего начала при формировании первичных лимфоидных фолликулов.
Реализация этих процессов в лимфатических узлах и пейеровых бляшках отличается деталями, тогда как формирование единичных фолликулов и фолликулярных структур слизистой оболочки носоглотки происходит по иным, в недостаточной степени изученным законам. Принципальное различие между этими группами лимфоидных структур состоит в том, что вторичные лимфоидные органы в основном формируются в эмбриогенезе, тогда как лимфоидные образования слизистых оболочек могут появляться в постнатальном периоде, в частности, под влиянием микрофлоры.
У взрослых организмов способность фолликулярных дендритных клеток к выработке хемокинов и факторов выживания поддерживается благодаря контактам с В-лимфоцитами, стимулирующими эту способность у дендритных клеток. Таким образом, В-лимфоциты в определенной степени сами формируют свое микроокружение. У мышей, дефектных по генам лимфотоксинов или их рецепторов, не развиваются фолликулярные дендритные клетки и не формируются лимфоидные фолликулы, а при определенной комбинации дефектов не образуются лимфатические узлы.
Т-клетки в лимфатических узлах располагаются в паракортикальных зонах, находящихся в глубоких слоях коры. Они окружают посткапиллярные венулы, через которые в ткань лимфатичекого узла из кровотока мигрируют лимфоциты. В отсутствие тимуса (при генетических дефектах или после тимэктомии сразу после рождения) эти зоны не формируются. Паракортикальные зоны служат нишами для наивных Т-лимфоцитов: стромальные клетки обладают способностью привлекать их, секретируя хемокины CCL19 (ELC) и CCL21 (SLC), к которым Т-клетки имеют рецептор (CCR7). Жизнеспособность мигрировавших Т-лимфоцитов поддерживается IL-7 — одним из главных факторов выживаемости наивных Т-клеток. Помимо соединительнотканных клеток, в формировании ниши для Т-лим- фоцитов участвуют дендритные клетки (истинные, происходящие из костного мозга). Поскольку эти дендритные клетки обычно охватывают своими отростками Т-лимфоциты, их называют интердигитальными (межпальцевыми) клетками. Интердигитальные клетки также продуцируют CCL19 и CCL21. TCR распознают на поверхности интердигитальных дендритных клеток молекулы MHC, несущие аутологичные пептиды. Это также повышает выживаемость наивных Т-лимфоцитов (см. ниже). При распознавани в составе этих молекулярных комплексов чужеродных пептидов, Т-клетки активируются.
Пространство между фолликулами и вне паракортикальных зон занято смешанной популяцией лимфоцитов, включающей как Т-, так и В-лимфо- циты. Здесь эти клетки контактируют между собой, что важно для развития иммунного ответа. Лимфоциты пребывают в лимфатических узлах временно. Они постоянно рециркулируют. Соотношение CD4+ и CD8+ Т-клеток, а также других субпопуляций Т-лимфоцитов в лимфатических узлах примерно соответствует таковому в крови (см. табл. 3.8).
Мозговая зона лимфатических узлов содержит мозговые шнуры, между которыми расположены медуллярные синусы. В шнурах содержатся лимфоциты обоих классов. В ходе иммунного ответа здесь сосредоточивается значительная часть плазматических клеток. В синусах расположена лимфа, содержащая лимфоциты. Эта лимфа оттекает по эфферентным сосудам и впадает в кровеносную систему.
Структура лимфатических узлов схематически изображена на рис. 3.68. Со стороны выпуклой части лимфоузла через трабекулы в орган входят лимфатические сосуды, приносящие лимфу из дренируемых участков тела. В своей вогнутой части узел имеет ворота, через которые входят афферентные артерии и выходят эфферентные лимфатические сосуды и вены. Лимфатический узел окружен соединительнотканной капсулой, от которой внутрь отходят тонкие перегородки — трабекулы. Ткань лимфатического узла состоит из коры и мозгового вещества. В наружной части коры есть округлые образования — лимфоидные фолликулы, служащие местом сосредоточения В-лимфоцитов. В покоящихся лимфоузлах они имеют равномерную структуру, и их обозначают как первичные фолликулы. При иммунном ответе в фолликул, помимо В-лимфоцитов, мигрируют Т-клетки, в нем формируются центры размножения — зародышевые центры. Такой фолликул называют вторичным. Ключевую роль в формировании и функционировании фолликулов и развивающихся в них зародышевых центров играют фолликулярные дендритные клетки, по происхождению не имеющие отношения к обычным дендритным клеткам и сходные
Рис. 3.68. Строение лимфатического узла и локализация в нем Т- и В-лимфоцитов
(по Janeway С.А. et al., 2005)
с ними только по морфологии. Эти клетки имеют местное мезенхимальное происхождение.
Формирование Т- и В-зон лимфатических узлов, как и других вторичных лимфоидных органов, а также мозгового слоя тимуса определяется взаимодействием лимфоидных клеток и предшественников стромальных клеток, осуществляемое в эмбриогенезе. Ключевая роль в нем принадлежит мембранным лимфотоксинам (LT) — цитокинам семейства TNF — и их рецепторам. Из костного мозга в зачаток лимфатических узлов и других лимфоидных органов мигрируют специальные лимфоидные клетки, обозначаемые как клетки-индукторы лимфоидной ткани — LTIC (Lymphoid tissue inducing cells). Их мембранный фенотип — CD4+ CD3- CD45+ а4Р7+ IL-7Ra+ LTaP+ CXCR5+. LTIC экспрессируют дифференцировочный фактор RORyt (Retinoid-acid-receptor-related orphan receptor — орфановый рецептор, родственный рецептору ретиноидной кислоты) и транскрипционный фактор Id2. Судя по мембранным молекулам, эти клетки имеют костномозговое лимфоидное происхождение, но не относятся к зрелым субпопуляциям лимфоцитов. LTIC содержат на своей поверхности мембранную форму LTaP — тример, включающий 2 трансмембранные молекулы LTp и 1 молекулу ЕГа, расположенную внеклеточно. Другая сигнальная молекула LTIC, также относящаяся к семейству TNF, — RANKL (Recerptor activator of NFKB-ligand). В образовании лимфоидных органов участвуют также клетки-инициаторы лимфоидной ткани — LTIN, которые имеют фенотип CD4- CD3- IL-7Ra- CD11b+ CD11c+, свидетельствующий об их родстве с дендритными клетками. С этими двумя типами клеток взаимодействуют предшественники стромальных клеток — клетки-организаторы лимфоидной ткани — Lto.
LTp и RANKL распознаются рецепторами семейства TNFR на стромальных клетках, в результате чего в них генерируется сигнал, вызывающий экспрессию генов IL-7, хемокинов (CXCL13, CCL19, CCL21) и фактора TRANCE (TNF-related activation-induced cytokine). Все эти факторы действуют на лимфоидные клетки; кроме того, они привлекают зрелые лимфоциты, обеспечивают их жизнеспособность и способствуют их кластеризации с образованием зачатка фолликула. Затем мигрирующие в зачаток В-, Т- и NK-лимфоциты разделяются пространственно. К этому моменту в формирующемся органе появляются дендритные клетки, играющие роль организующего начала при формировании первичных лимфоидных фолликулов.
Реализация этих процессов в лимфатических узлах и пейеровых бляшках отличается деталями, тогда как формирование единичных фолликулов и фолликулярных структур слизистой оболочки носоглотки происходит по иным, в недостаточной степени изученным законам. Принципальное различие между этими группами лимфоидных структур состоит в том, что вторичные лимфоидные органы в основном формируются в эмбриогенезе, тогда как лимфоидные образования слизистых оболочек могут появляться в постнатальном периоде, в частности, под влиянием микрофлоры.
У взрослых организмов способность фолликулярных дендритных клеток к выработке хемокинов и факторов выживания поддерживается благодаря контактам с В-лимфоцитами, стимулирующими эту способность у дендритных клеток. Таким образом, В-лимфоциты в определенной степени сами формируют свое микроокружение. У мышей, дефектных по генам лимфотоксинов или их рецепторов, не развиваются фолликулярные дендритные клетки и не формируются лимфоидные фолликулы, а при определенной комбинации дефектов не образуются лимфатические узлы.
Т-клетки в лимфатических узлах располагаются в паракортикальных зонах, находящихся в глубоких слоях коры. Они окружают посткапиллярные венулы, через которые в ткань лимфатичекого узла из кровотока мигрируют лимфоциты. В отсутствие тимуса (при генетических дефектах или после тимэктомии сразу после рождения) эти зоны не формируются. Паракортикальные зоны служат нишами для наивных Т-лимфоцитов: стромальные клетки обладают способностью привлекать их, секретируя хемокины CCL19 (ELC) и CCL21 (SLC), к которым Т-клетки имеют рецептор (CCR7). Жизнеспособность мигрировавших Т-лимфоцитов поддерживается IL-7 — одним из главных факторов выживаемости наивных Т-клеток. Помимо соединительнотканных клеток, в формировании ниши для Т-лим- фоцитов участвуют дендритные клетки (истинные, происходящие из костного мозга). Поскольку эти дендритные клетки обычно охватывают своими отростками Т-лимфоциты, их называют интердигитальными (межпальцевыми) клетками. Интердигитальные клетки также продуцируют CCL19 и CCL21. TCR распознают на поверхности интердигитальных дендритных клеток молекулы MHC, несущие аутологичные пептиды. Это также повышает выживаемость наивных Т-лимфоцитов (см. ниже). При распознавани в составе этих молекулярных комплексов чужеродных пептидов, Т-клетки активируются.
Пространство между фолликулами и вне паракортикальных зон занято смешанной популяцией лимфоцитов, включающей как Т-, так и В-лимфо- циты. Здесь эти клетки контактируют между собой, что важно для развития иммунного ответа. Лимфоциты пребывают в лимфатических узлах временно. Они постоянно рециркулируют. Соотношение CD4+ и CD8+ Т-клеток, а также других субпопуляций Т-лимфоцитов в лимфатических узлах примерно соответствует таковому в крови (см. табл. 3.8).
Мозговая зона лимфатических узлов содержит мозговые шнуры, между которыми расположены медуллярные синусы. В шнурах содержатся лимфоциты обоих классов. В ходе иммунного ответа здесь сосредоточивается значительная часть плазматических клеток. В синусах расположена лимфа, содержащая лимфоциты. Эта лимфа оттекает по эфферентным сосудам и впадает в кровеносную систему.