
Лимфоидные органы — динамичные образования: лимфоциты непрерывно перемещаются в стромальном каркасе, покидают лимфоидные органы и вновь возвращаются в них. Этот круговорот лимфоцитов обозначают термином «рециркуляция». В процессе рециркуляции лимфоциты покидают лимфоидные органы с эфферентной лимфой (из селезенки они выходят с кровью). Через грудной лимфатический проток лимфоциты проникают в кровяное русло. Затем через посткапиллярные венулы клетки вновь проникают в лимфоидный орган (тот же или другой), внутри которого перемещаются в специфические места их локализации. Затем клетки вновь покидают орган и выходят в циркуляторное русло и т.д. В нелимфоидных органах лимфоциты практически не циркулируют.
Из сказанного следует, что выявляемые в крови лимфоциты находятся в ней недолго (примерно 30 мин), но в течение суток многократно (4—5 раз) покидают ее и вновь возвращаются. Таким образом, несмотря на то, что в крови лимфоциты составляют всего около 0,5% от общего числа клеток, их состояние достаточно точно отражает состояние популяции лимфоцитов в целом организме. Это очень существенно, поскольку именно лимфоциты крови человека служат наиболее обычным и доступным объектом изучения как в экспериментальных, так и в клинико-иммунологических исследованиях. Субпопуляционный состав лимфоцитов крови человека представлен в табл. 3.23.
Таблица 3.23. Нормальные показатели содержания в крови взрослых людей основных популяций и субпопуляций лимфоцитов (по С.В. Хайдукову, 2008)
|
Клетки |
Относительное содержание*, % |
Число клеток в 1 л (М+m) |
|
Лимфоциты (CD45+)** |
32+4 |
1,363—2,808х109 |
|
В-клетки ^D3-CD19+) |
12+5 |
0,111—0,376х109 |
|
Т-клетки (CD3+CD19-) |
73+12 |
0,946-2,079х109 |
|
арТ-клетки (CD3+apTCR+) |
70,5+9,7 |
0,022—0,115х109 |
|
yST-клетки (CD3+ySTCR+) |
4,6+2,8 |
0,924—1,964х109 |
|
Т-хелперы (CD3+CD4+) |
45+10 |
0,576—1,336х109 |
|
Т-киллеры (CD3+CD8+) |
27+8 |
0,372—0,974х109 |
|
Регулятоные T-клетки (CD3+CD4+CD25hi) |
3,7+2,05 |
0,009—0,078х 109 |
|
NKT-клетки (CD3+CD56+CD16+) |
3+3 |
0,007—0,165х109 |
|
Т-хелперы наивные (CD3+CD4+CD45RA+) |
30+10 |
0,272—1,123х109 |
|
Т-хелперы активированные/памяти (CD3+CD4+CD45R0+CD29+) |
15+10 |
0,068—0,702х109 |
|
NK-клетки (CD45+CD56+СD16+) |
13+5 |
0,123—0,369х109 |
* Указано среднее арифметическое со стандартной ошибкой.
** Относительные и абсолютные количества лимфоцитов определяли от общего количества лейкоцитов.
Относительные и абсолютные количества субпопуляций Т-, В- и NK-клеток определяли от общего количества лимфоцитов.
Способность клеток находить «свое место» в организме называют хомингом. Существует две группы механизмов, задействованных в данном процессе, — контактные механизмы, обеспечиваемые молекулами адгезии, находящимися на поверхности лимфоцитов, и хемотаксис, определяющий направленность движения клеток, зависящую от наличия на их поверхности хемокиновых рецепторов.
Попав в сосудистую сеть лимфоидного органа, лимфоциты реагируют на хемокины, вырабатываемые клетками эндотелия в специализированном для миграции лимфоцитов отделе сосудов — посткапиллярных венулах.
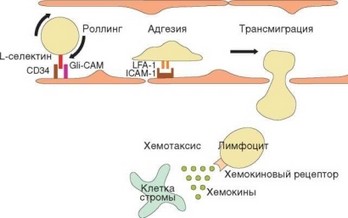
Рис. 3.74. Схема транссосудистой миграции лимфоцитов. Отражены основные стадии эмиграции лимфоцитов через высокий эндотелий посткапиллярных венул лимфоидных органов. Указаны молекулы адгезии, важные для осуществления этого процесса; отражена роль хемокинов в привлечении клеток
Только в посткапиллярных венулах лимфоидных (но не других) органов есть высокий эндотелий, образованный спонтанно активированными клетками (аналогичный высокий эндотелий может быть индуцирован в других органах только под влиянием провоспалительных факторов). Клетки этого эндотелия секретируют р-хемокин ССЫ9, распознаваемый рецептором CCR7. Этот рецептор экспрессируют на поверхности наивные лимфоциты как Т-, так и В-класса. Таким образом, указанные эндотелиальные клетки способны привлекать как Т-, так и В-лимфоциты.
Далее происходит трансмиграция лимфоцитов по механизму, аналогичному миграции лейкоцитов в очаг воспаления (отличия касаются конкретных молекул адгезии, определяющих направление миграции — рис. 3.74). Ключевую роль при этом играет начальный момент миграции, обусловленный взаимодействием мембранного L-cелектина (СD62L) лимфоцитов с адрессинами (GlyCAM, CD34, PSGL-1) эндотелиальных клеток. Именно наличие CD62L на клеточной мембране определяет способность наивных лимфоцитов мигрировать в лимфатические узлы и пейеровы бляшки, в связи с чем CD62L называют рецептором хоминга. Это слабое взаимодействие обусловливает феномен качения (перекатывания лимфоцита вдоль сосудистой стенки). Вслед за этим устанавливается более прочное взаимодействие между р2-интегрином лимфоцитов LFA-1 и его рецептором на эндотелиальной клетке — ICAM-1. Взаимодействие усиливается благодаря «активации» р2-интегрина (т.е. изменения конформации молекулы, в результате которого повышается его сродство к рецептору — см. раздел 2.3.1.2). Это обусловливает прочное прилипание лимфоцита к сосудистой стенке. Затем под влиянием хемотаксических сигналов, поступающих из ткани
лимфоидного органа, лимфоцит начинает мигрировать из сосуда между эндотелиальными клетками.
Вышедшие из сосуда клетки попадают в примыкающие к посткапиллярным венулам тимусзависимые зоны, стромальные и дендритные клетки которых секретируют хемокины CCL19 (ELC) и С^21 (SLC), распознаваемые рецептором CCR7 на лимфоцитах. До этого момента Т- и В-клетки перемещаются в одном и том же направлении. После того как лимфоцит попадает во внутреннюю среду органа, их пути расходятся. Т-клетки остаются в тимусзависимой зоне (в паракортексе лимфатических узлов, параартериальной муфте белой пульпы селезенки и Т-зонах пейеровых бляшек). В-клетки реагируют на хемокин CXCR13 (BLC), к которому они экспрессирут рецептор СХСR5. Этот хемокин секретируют стромальные клетки фолликулов, что и определяет направление миграции В-лимфоцитов. В-лимфоциты, не несущие СХСR5, не поступают в фолликулы и локализуются в прилежащей к фолликулу зоне. Механизмы, определяющие дальнейшее перемещение лимфоцитов из органа в эфферентную лимфу, пока не установлены.
В процессе рециркуляции отсутствует «привязка» хоминга к конкретному лимфатическому узлу или пейеровой бляшке. Так, лимфоцит, покинувший шейный лимфоузел, может затем проникнуть, например, в паховый лимфатический узел или в пейерову бляшку. Принципиально лишь то, что клетки не выходят из системы вторичных лимфоидных органов. Степень специфичности хоминга минимальна в селезенке, так как лимфоциты проникают в белую пульпу при излиянии крови в краевой зоне. Только последующее перемещение в Т- и В-зоны осуществляется по описанному выше механизму.
Все сказанное выше относится к наивным лимфоцитам. После активации лимфоцитов антигеном экспрессия молекул адгезии и хемокиновых рецепторов на их поверхности существенно изменяется. Это определяет иные пути миграции клеток после контакта с антигеном. Активированные лимфоциты и клетки памяти очень слабо экспрессируют L-селектин, что обусловливает низкую вероятность их попадания в лимфатические узлы. В то же время они приобретают способность мигрировать в очаги воспаления, расположенные в нелимфоидных органах, а также в барьерные ткани (рис. 3.75). Пути миграции активированных лимфоцитов и клеток

Рис. 3.75. Мембранный фенотип Т-клеток определяет пути их миграции. Указаны молекулы адгезии, определяющие различные направления миграции Т-клеток
памяти будут специально рассмотрены далее (см. раздел 3.6.5.4). Благодаря различной экспрессии рецепторов хоминга в разных тканях формируется несколько относительно автономных путей рециркуляции. Пути рециркуляции наивных лимфоцитов и клеток памяти могут пересекаться. Так, и наивные клетки, и клетки памяти, активированные в кишечнике, попадают в брыжеечные лимфатические узлы.
Интенсивность рециркуляции Т-лимфоцтов существенно выше, чем В-лимфоцитов. У человека через каждый лимфатический узел за сутки проходит 0,3х10п, через селезенку — 2,5х10п, а через кровь — 5х10п лимфоцитов. Длительность пребывания Т-клеток в кровотоке за 1 цикл рециркуляции — около 30 мин. Благодаря рециркуляции происходит постоянное «перемешивание» лимфоидных клеток с сохранением описанного выше порядка: наивные клетки, как правило, не покидают лимфоидной ткани, тогда как клетки памяти, наоборот, часто выходят за ее пределы, мигрируя, например, в эпителиальные органы и ткани, а также в очаги воспаления. Внутри лимфоидных органов клетки также занимают соответствующие места. Упорядоченность локализации лимфоцитов различных типов очень важна для развития иммунного ответа.