
Тимус, или вилочковая железа, расположен за рукояткой грудины. У человека и большинства млекопитающих он состоит из двух долей, у птиц и некоторых млекопитающих число долей значительно больше и варьирует.
Долгое время функция тимуса была неизвестна, поскольку его удаление у взрослых животных не приводило к серьезным последствиям. Только в 1961 г. австралийский иммунолог Дж. Миллер (J.F.A.P. Miller) установил, что удаление тимуса у новорожденных мышей приводит к нарушению развития лимфоцитов — не всех, но большинства. Так был установлен факт гетерогенности лимфоцитов и была обнаружена популяция тимусзависимых или Т-лимфоцитов, т.е. лимфоцитов, развитие которых происходит в тимусе.
Структура тимуса
Тимус состоит из двух основных частей — коры и мозгового вещества. В коре выделяют наружный, субкапсулярный слой, глубокую кору и кортикомедуллярную зону (или кортикомедуллярное сочленение). Тимус покрыт соединительнотканной капсулой, от которой внутрь коры отходят междольковые перегородки, разделяющие кору на дольки. Мозговая часть тимуса не разделена на дольки. Кортикомедуллярное сочленение служит воротами тимуса, в которые входят и выходят кровеносные сосуды и нервы. Сосудистая сеть тимуса бедна («бледный орган»). Наиболее важны в функциональном отношении посткапиллярные венулы, через стенку которых в тимус проникают клетки-предшественники и выходят зрелые Т-лимфоци- ты. Схематически строение тимуса отражено на рис. 3.57.
Гистологически тимус представляет лимфоэпителиальный орган. Он имеет трехслойную структуру, включающую обычную соединительнотканную строму, эпителиальный ретикулум и лимфоидную составляющую (табл. 3.17). Эпителиальные и большая часть соединительнотканных элементов тимуса имеют местное происхождение. Лимфоциты тимуса,
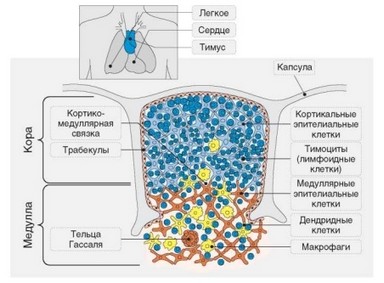
Рис. 3.57. Схема строения тимуса (по Janeway C.A., 2005)
называемые тимоцитами, а также макрофаги, дендритные и тучные клетки происходят из клеток-предшественников, мигрирующих в этот орган из костного мозга. Тимоциты численно преобладают и являются транзиторны- ми элементами, поскольку подавляющее большинство их покидает тимус по мере созревания.
Соединительнотканная строма формирует капсулу, междольковые перегородки и периваскулярное пространство. В последнем содержатся фибробласты, соединительнотканные волокна, макрофаги, тучные клетки, В-лимфоциты, нейтрофилы. Доля соединительнотканного пространства в тимусе очень невелика. Основной объем органа занимает эпителиальное (внутреннее) пространство — участки тимуса, ограниченные эпителиальными клетками и базальной мембраной. Эпителиальные клетки образуют трехмерный каркас — эпителиальный ретикулум, внутри которого происходят основные события, связанные с развитием Т-лимфоцитов. Таким образом эпителиальная составляющая тимуса фактически представляет паренхиму органа. В эпителиальном каркасе кортикомедуллярной зоны содержатся макрофаги, а в мозговом веществе — еще и дендритные клетки.
Специфическая функция тимуса состоит в обеспечении развития (созревания, селекции, дифференцировки) Т-лимфоцитов. В этом задействованы преимущественно эпителиальные клетки. Определенный вклад в этот процесс вносят соединительнотканные элементы и дендритные клетки тимуса. Доказательства различного происхождения лимфоидных и эпителиальных клеток тимуса, а также данные о гистогенезе тимусного эпителия были получены в опытах с межвидовой трансплантацией клеток эмбрионов кур
и перепелов, а также с культивированием и пересадкой меченых клеток «закладки» тимуса.
Гистогенез тимусного эпителия
Эпителиальные клетки тимуса происходят из энтодермы 3—4-го глоточного кармана (существовавшее до недавнего времени представление о смешанном, энтодермально-эктодермальном происхождении тимусного эпителия отвергнуто). В их развитии участвует нервный гребень, служащий источником индукторов дифференцировки эпителия. Кроме того, клетки, происходящие из мезенхимы нервного гребня, мигрируют в зачаток тимуса (некоторые считают, что из них развиваются клетки, продуцирующие нейрогормоны). Важным достижением было обнаружение общих клеток- предшественников кортикального и медуллярного эпителия. Подсадка этих клеток под почечную капсулу приводит к развитию эктопического тимусного эпителия.
Таблица 3.17. Нелимфоидные клетки тимуса
|
Тип клеток |
Разновид ности |
Локали зация |
Маркеры |
Продукты |
Функции |
|
Эпителиальные клетки |
Субкапсу лярные |
Субкапсулярная зона коры |
СК5- СК8+; MHC-II, LFA-3, ICAM-1, VCAM-1 |
Гормоны тимуса: тимулин, а1-тимо- зин, тимопоэтин. Цитокины: IL-1a, IL-1P, TNFa, IL-6, IL-7, LIF, IL-8, CXCL9, CXCL10, CXCL12, CCL1, CCL2, CCL5 CCL11, CCL17, CCL19, CCL21, CCL22, CCL25 |
Дифферен- цировка тимо- цитов |
|
Кортикальные, в том числе клетки- няньки |
Кора тимуса |
СК5- СК8+; MHC-II, LFA-3, ICAM-1, VCAM-1 |
Цитокины: IL-1p, IL-6, IL-7, GM-CSF, SCF, СХСЬ12, CCL25 |
Дифферен- цировка тимо- цитов, положительная селекция |
|
|
Медул лярные |
Мозговой слой |
СК5+СК8-; МНС-II, СD80, CD86, ICAM-1, VCAM-1, LFA-3, CD40 |
Гормоны тимуса: тимулин, а1-тимо- зин, тимопоэтин. Цитокины: IL-1a, IL-1P, TNFa, IL-6, IL-7, LIF, IL-8, CXCL9, CXCL10, CXCL12, CCL1, CCL2, CCL5 CCL11, CCL17, CCL19, CCL21, CCL22, CCL25 |
Дифферен- цировка тимо- цитов, отрицательная селекция |
|
|
Клетки телец Гассаля |
Мозговой слой |
CK5+CK8-; MHC-II |
TSLA |
Дифферен- цировка регуляторных T-клеток |
Окончание табл. 3.17
|
Тип клеток |
Разновид ности |
Локали зация |
Маркеры |
Продукты |
Функции |
|
Миелоидные клетки |
Дендритные клетки (лимфоидные и миело- идные) |
Мозговой слой |
MHC-II, CD80, CD86, CD40, TLR, CD1k |
Миелоидные — IFNa, IL-12, CCL10, CCL17, CCL 25, лимфоидные — CCL4, CCL10, M-CSF |
Отрицательная селекция тимоцитов, дифференци- ровка |
|
Макро фаги |
Кортико- медуллярный и мозговой слои |
CD14, IFNyR, VLA-4, CD40, TLR |
IL-1, TNF, IL-6, про- стагландин E2 |
Дифферен- цировка тимоцитов, источник цитокинов |
|
|
Миоидные клетки |
Мозговой слой |
AcChR |
TNFa, IL-8, IL-6 |
Пролиферация, диф- ференцирвока тимоцитов |
|
|
Фибробласты |
Внеэпите лиальный компарт- мент |
ICAM-1 |
SCF, IL-7, GM-CSF, Flt-3-L, LIF, IL-6, IL-1, FGF, простаглан- дин G, простаглан- дин F , TFGp, CXCL12 |
Механическая функция, источник цитокинов |
|
Формирование и развитие закладок тимуса и локализующейся рядом закладки паращитовидных желез контролируется каскадом дифференциро- вочных факторов (Hoxa3, Pax1, Pax9, Eya1 и Six1), экспрессируемых в глоточной энтодерме (рис. 3.58). Наиболее важную роль играют факторы Hoxa3 и Рах9, отсутствие которых приводит к атимии. Фактору Hoxa3 приписываются роль позиционирующего агента, который определяет связь закладки рудиментов тимуса и паращитовидных желез с 3-м глоточным карманом, а фактор Рах9 отвечает за обособление закладки тимуса от глотки. Разделение закладок тимуса и паращитовидных желез и начало дифференцировки эпителиальных клеток тимуса связывают с другими транскрипционными факторами — Foxn1 и Gcm2. Наконец, очередной этап развития эпителиальной закладки тимуса и ее функциональная полноценность контролируются фактором Foxn1 (Foxhead box N1); мутации кодирующего его гена служат основой патологии тимуса у мышей nude («голые» мыши), у которых эпителиальная строма тимуса формируется, но не функционирует, поскольку не заселяется лимфоидными предшественниками. Экспрессию гена Foxn1 регистрируют в клетках глоточной энтодермы в срок Е.10,5 (буква Е означает эмбриональное развитие, цифра — сутки).
Родоначальные клетки тимусного эпителия впервые выявляются на сроке Е.11,5. Они одновременно экспрессируют цитокератины (CK) 5 и 8. Через сутки (Е.12,5) происходит миграция первых лимфоидных клеток-предшест- венников в закладку тимуса. Осуществляется она не через сосуды, которые пока отсутствуют, а путем прямого перемещения в тканях. На протяжении трех следующих суток (до Е.15,5) происходит формирование функциональ-

Рис. 3.58. Формирование и развитие эпителия тимуса. Отражена дифференцировка эпителиальных клеток тимуса с указанием экспрессируемых ими цитокератинов и дифференцировочных факторов. Указаны основные события раннего развития закладки тимуса
но полноценного эпителиального ретикулума — создается его трехмерная структура, происходит васкуляризация и дифференцировка родоначальных эпителиальных клеток тимуса в кортикальные (СК8+) и медуллярные (СК5+) эпителиальные клетки. В этот период на развитие тимусного эпителия влияют мезенхимальные производные нервного гребня через продукты генов Fgf (Fibroblast growth factors), Wnt (Wingless), Bmp (Bone morphogenetic proteins) и Shh (Sonic-hodgehog homologue). Наиболее полно изучена роль ростового фактора FGF (продуктов генов Fgf), секретируемого клетками экто/мезодермы. На ранних этапах развития тимуса функционируют факторы FGF-7 и FGF-10. На эпителиальных клетках тимуса экпрессирован рецептор для этих факторов — FgfR-IIIb. Продукт гена Shh регулирует экспрессию фактора Tbx1 в эпителиальных клетках тимуса; дефект этого фактора у человека проявляется как синдром Ди Джоджи (дисгенезия тимуса).
В более поздний период очень важную роль приобретает взаимодействие между эпителиальными и лимфоидными клетками, в реализации которого основную роль играют продукты гена Wnt. Важнейше события последующего развития эпителиальной стромы:
- разделение на кору и мозговой слой с формированием кортико-медуллярной структуры органа (между 13-14-ми сутками);
- экспрессия на эпителиальных клетках функционально важных молекул.
В первую очередь следует отметить экспрессию молекул Jagged и Delta — лигандов основных дифференцировочных факторов Т-клеток — Notch-1 и Notch-3. Не менее важное свойство эптелиальных клеток тимуса —
секреция IL-7 как основного лимфопоэтического цитокина, служащего фактором выживаемости и роста развивающихся тимоцитов. Несколько позже эпителиальные клетки тимуса начинают экспрессировать молекулы MHC-II, необходимые для осуществления дифференцировки и селекции клонов Т-клеток. Эти маркеры сначала экспрессируются на эпителиальных клетках мозгового слоя (E.14), а затем коркового (E.16). Формирование структуры мозгового слоя происходит при активном участии медуллярных тимоцитов и специализированных клеток, участвующих в гистогенезе вторичных лимфоидных органов (см. далее) — LTIC (Lymphoid tissue inducer cells). Экспрессируемые ими мембранные рецепторы семейства TNF — гетеродимер лимфотоксинов LTaP2 и RANKL (Receptor activator of nuclear factor NF-кВ ligand) — взаимодействуют с соответствующими рецепторами семейства TNFR на поверхности эпителиальной клетки. В результате клетки эпителия начинают вырабатывать комплекс хемокинов и цитокинов, обеспечивающих развитие и структурную организацию мозгового слоя и поздние этапы дифференцировки тимоцитов.
Заселение тимуса лимфоидными клетками и гематотимический барьер
Заселение тимуса кроветворными клетками-предшественниками происходит импульсно. Так, у птиц четко выявляют 3 периода заселения, между которыми происходит развитие тимоцитов и их эмиграция на периферию. У мышей выявлено 2 периода заселения — 11—13-е и 17—19-е сутки эмбрионального развития. Клетки-предшественники, мигрирующие из фетальной печени, созревают в у5Т-клетки с ограниченной вариабельностью TCR, покидающие тимус еще в эмбриональном периоде. Первая волна эмиграции происходит на 15—16-е сутки внутриутробного развития; сначала эмигрирующие у5Т-клетки поступают в кожу, затем в слизистые оболочки языка и репродуктивной системы самок. В последующем эти клетки самоподдерживаются местно. Позже в тимусе начинают формироваться у5Т-клетки с более широким спектром специфичностей TCR, покидающие тимус вскоре после рождения (до 13 сут) и заселяющие различные слизистые оболочки. С 16-х суток эмбриогенеза в тимусе начинают развиваться apT-клетки. На 17—19-е сутки внутриутробного развития происходит вторая волна заселения тимуса клетками-предшественниками. Считают, что потомки клеток-предшественников, поступивших в тимус во время этой волны, не покидают его, образуя резерв для срочной регенерации органа после стресса и воздействия разнообразных повреждающих факторов. У человека ранние этапы функционирования тимуса происходят принципиально так же, как у мышей. Закладка эпителия тимуса происходит на 6-й неделе развития плода, первая волна заселения осуществляется на 8-й неделе эмбрионального развития.
После рождения устанавливается режим миграции клеток через тимус, характерный для взрослых животных. Клетки-предшественники из костного мозга поступают в тимус постоянно в очень небольшом количестве (в тимусе мыши есть всего около 200 «ниш», способных принять мигрирующие клетки), а зрелые клетки из тимуса эмигрируют постоянно. Миграция клеток-предшественников в тимус представляет довольно сложный процесс. Прежде всего тимус должен поставлять хемотаксические сигналы.

Рис. 3.59. Гематотимический барьер и его преодоление предшественниками тимо- цитов: 1 — сосудистой стенки; 2 — внеэпителиального пространства; 3 — базальной мембраны тимусного эпителия
Передача таких сигналов происходит при помощи CXCL12 (SDF-1), С^25 (TECK) и некоторых других хемокинов, продуцируемых эпителиальными клетками тимуса. Для восприятия таких сигналов на поверхности клеток- предшественников имеются рецепторы — СXCR4, ССR9 и ССR5. Блокада этих рецепторов антителами затрудняет заселение тимуса.
Тимус изолирован от проникновения в него клеток извне с помощью гематотимического барьера (рис. 3.59). Этот барьер состоит из трех компонентов: первый — эндотелий посткапиллярных венул. Мигрирующие клетки преодолевают его благодаря взаимному с клетками эндотелия сосудов распознаванию молекул адгезии. Установлена роль в иммиграции кле- ток-прешественников в тимус селектинов L и Р, а4-интегринов (VLA-4 или а4р1, а4р7). Рецепторы названных интегринов — фибронектин, VCAM-1 и MadCAM экспрессируются в зонах проникновения клеток-предшествен- ников в тимус. В этом процессе задействованы также р2-интегрин LFA-1, Pj-интегрин VLA-6 и их рецетпоры ICAM-1 и ламинин соответственно. Судя по блокирующему эффекту антител, важную роль при миграции клеток в тимус играют молекулы CD44 (экспрессируемые на поверхности мигрирующих клеток и исчезающие вскоре после поступления в тимус), а также молекула VAP-1 (Vascular adhesion protein 1), экспрессируемая только в венулах тимуса. Второй компонент барьера образован макрофагами и другими клетками периваскулярного пространства. Избежать поглощен- ния макрофагами клеткам помогает блокада сиаловой кислотой свободных углеводных остатков их поверхностных гликопротеинов. Для перемещения в межклеточном пространстве важна экспрессия на поверхности клеток- предшественнков Pj-интегринов, в частности VLA-4 и VLA-6, взаимодействующих с белками межклеточного матрикса, — соответственно фибронек- тином и ламинином. Продвижению клеток-предшественников способствует их инвазивность, обусловленная выделением ферментов гиалуронидазы и коллагеназы, расплавляющих межклеточный матрикс. Третий и наименее
проницаемый компонент барьера — эпителиальные клетки, ограничивающие эпителиальное пространство тимуса, а также их базальная мембрана. Этот барьер преодолевается благодаря экспрессии на клетках-предшествен- никах Р-селектинов, взаимодействующих с молекулами GlyCAM, MadCAM, и молекулы CD44, реагирующей с гиалуронатом межклеточного матрикса. Гематотимический барьер распространяется на кору, но не на мозговое вещество тимуса, в которое могут мигрировать циркулирующие клетки крови. Полагают, что в преодолении барьера, образованного базальной мембраной, важная роль принадлежит металлопротеиназам.
Разнообразие эпителиальных клеток тимуса и их функции (см. табл. 3.17)
Эпителиальные клетки субкапсулярного, глубокого коркового и мозгового слоев тимуса отличаются рядом свойств. Все эти клетки имеют звездчатую форму. В тимусе молодых животных содержится много тимоцитов, поэтому эпителиальные клетки трудно выявить, поскольку они сдавлены тимоцитами. Изучение этих клеток проводят in vitro в культурах клеток. Функции эпителиальных клеток проявляются в полной мере только в трехмерной культуре — при выращивании клеток на губчатых каркасах. Выделяют секреторные и ретикулярные (поддерживающие) эпителиальные клетки тимуса, однако это деление условно, поскольку практически все клетки тимусного эпителия способны выполнять обе функции и преобладание той или другой из них зависит от локализации и функционального состояния клетки.
По-видимому, основные функции эпителиальных клеток тимуса реализуются в форме контактных взаимодействий. Выше было сказано о ключевой роли межклеточных взаимодействий, в основе которых лежит взаимное распознавание лигандов и рецепторов дифференцировочных факторов семейства Notch. Аналогично проявляют свое действие некоторые другие дифференцировочные факторы. Помимо этого большая группа мембранных молекул обеспечивает межклеточную адгезию, в частности, контакты эпителиальных и лимфоидных клеток тимуса. В формировании этих контактов основную роль играют молекулы адгезии — CD2 тимоцитов и LFA-3 (CD58) эпителиальных клеток. Важную дополнительную роль играют интегрины и их рецепторы. На поверхности эпителиальных клеток есть рецепторы рг и Р2-интегринов — соответственно, молекулы ICAМ-1 и VCAM-1, взаимодействующие с интегринами LFA-1 (Ргинтегрин), VLA-4 и VLA-6 (Р2-интегри- ны) на поверхности тимоцитов. Достаточно прочные контакты между эпителиальными и лимфоидными клетками важны как для передачи ростовых и дифференцировочных сигналов, так и для осуществления селекции. Как уже отмечалось выше, кортикальные эпителиальные клетки отвечают за положительную, а медуллярные — за отрицательную селекцию. И в том, и в другом случае необходима экспрессия на эпителиальных клетках молекул MHC-I и MHC-II. В тимусе многие эпителиальные клетки, особенно кортикальные, экспрессируют на поверхности эти молекулы. MHC-II в трехмерной культуре эпителиальных клеток тимуса экспрессируются сильнее, чем в обычной двумерной культуре, что, как полагают, и определяет разную степень функциональной активности эпителиальных клеток в указанных типах культур. Эпителиальные клетки тимуса несут на своей поверхности молекулы, необходимые для осуществления эффективной передачи сигнала в процессе селекции. Экспрессия костимулирующих молекул CD80, CD86 и CD40 закономерно проявляется только на эпителиальных клетках медуллярной зоны тимуса.
Среди эпителиальных клеток тимуса выделяют особую их разновидность — клетки-няньки. Они расположены в слоях коры, промежуточных между поверхностными и глубокими. Эти клетки образуют агрегаты путем обволакивания тимоцитов (сначала полагали, что клетки-няньки поглощают тимоциты). Одна клетка-нянька может окружать несколько сотен тимоцитов. Через 10—12 сут после выделения и культивирования таких агломератов in vitro происходит выход из них тимоцитов. Считалось, что таким образом формируется микроокружение, оптимальное для дифферен- цировки Т-клеток. Однако после обнаружения апоптоза тимоцитов внутри клеток-нянек возникло предположение об участии этой формы эпителиальных клеток в элиминации тимоцитов, не поддержанных селекцией.
Своеобразную морфологическую структуру, образованную эпителиальными клетками, представляют тельца Гассаля. Они расположены в медуллярной части тимуса человека и некоторых животных (их нет в тимусе мыши) и выглядят как многослойное чешуйчатое образование, напоминающее срез луковицы. Эпителиальные клетки телец Гассаля несколько отличаются по мембранным маркерам и гуморальным продуктам от других медуллярных клеток. В центре телец Гассаля расположены нежизнеспособные ороговевшие клетки, что послужило основанием считать тельца Гассаля участками ороговения эпителиальных клеток. Однако недавно установлено, что клетки телец Гассаля секретируют цитокин TSLP, влияющий на дендритные клетки. Дендритные клетки, активированные TSLP, играют важную роль при дифференцировке регуляторных Т-клеток; кроме того, они индуцируют дифференцировку аллергического варианта ТИ2-кле- ток (см. раздел 4.5.1.3).
Функция эпителильных клеток тимуса реализуется также через выделяемые ими гуморальные факторы — прежде всего цитокины. Важнейший продукт эпителиальных клеток тимуса — IL-7. Образуемый в тимусе IL-7, пополняет (наряду с IL-7, секретируемым стромальными клетками периферических лимфоидных органов) пул фактора в периферической части иммунной системы, необходимый для поддержания гомеостаза популяции Т-лимфоцитов (см. раздел 3.4.3). Эпителиальные клетки тимуса секретиру- ют цитокины, обычно выступающие в качестве провоспалительных факторов — IL-ip, IL-6, IL-8, TNFa. Однако в тимусе они выполняют функцию дифференцировочных факторов, нередко оказывая действие в комбинации друг с другом. По-видимому, цитокины тимуса формируют «малую цито- киновую сеть», обладающую автономией по отношению к общей цитоки- новой сети организма. В этой малой сети некоторые цитокины (например, воспалительные) могут выполнять функции, вне тимуса не являющиеся для них основными. Особую роль в тимусе играют хемокины, определяющие направление миграции тимоцитов внутри органа, о чем будет упомянуто ниже. Наконец, клетки тимуса (преимущественно эпителиальные) секрети- руют многочисленные нейропептиды и пептидные гормоны, которые будут рассмотрены далее (см. раздел 3.4.1.4).
Лимфоидные клетки тимуса (см. табл. 3.13)
Тимоциты образуют наиболее многочисленную популяцию клеток тимуса. Здесь представлены все стадии развития тимоцитов, о которых говорилось выше (см. раздел 3.3.2.3 и рис. 3.45, 3.46). Наименее зрелые CD-CD8-CD3- тимоциты расположены в субкапсулярной зоне тимуса. На их долю приходится 3—5% от общего числа тимоцитов. На первой стадии (DN1) происходит чрезвычайно интенсивная пролиферация клеток. На следующих стадиях развития дваждыотрицательных клеток осуществляется перестройка V-генов TCR, экспрессия мембранных рецепторов и формирование определяемого ими первичного (т.е. не прошедшего селекции) антигенраспознающего репертуара. Все этапы развития тимоцитов проходят в определенных зонах тимуса с характерным микроокружением, формируемым в наибольшей степени эпителиальными клетками и их продуктами (рис. 3.60).
Клетки с мембранным фенотипом CD4+CD8+ составляют наиболее многочисленную фракцию тимоцитов. На их долю приходится 70—75% общего числа лимфоидных клеток тимуса. Они локализуются в коре (преимущественно в ее глубоких слоях). На этой стадии происходят оба этапа селекции — положительная и отрицательная, формируется вторичный (селекционированный) антигенраспознающий репертуар, обеспечивающий распознавание пептидов в составе молекул MHC (рис. 3.61). Кортикальные тимоциты особенно чувствительны к индукции апоптоза в связи со слабой экспрессией антиапоптотических факторов. В частности, они погибают при действии глюкокортикоидов (кортизончувствительные тимоциты). В «отобранных» тимоцитах, получивших сигналы к дальнейшему развитию от эпителиальных клеток, усиливается экспрессия антиапоптотических
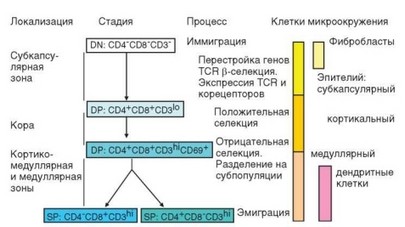
Рис. 3.60. Стадии развития тимоцитов и контролирующие их типы эпителия. Отражена связь основных этапов развития тимоцитов с различными клетками стромы и эпителиального ретикулума тимуса. Указаны зоны тимуса, в которых локализуются клетки (слева) и основные процессы развития тимоцитов (справа)
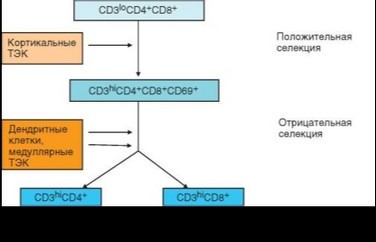
факторов. Одновременно возрастает экспрессия рецепторного комплекса TCR-CD3 и появляется маркер активации CD69.
В мозговом слое тимуса сосредоточены зрелые моноположительные тимоциты. На долю CD4+CD8- клеток приходится около 10% (из них 2—3% — регуляторные CD4+CD25+ клетки), на долю CD4-CD8+ клеток — около 5% лимфоидных клеток тимуса. Все эти клетки экспрессируют комплекс TCR—CD3 с высокой плотностью. Медуллярные тимоциты содержат так называемые Т-рецепторные эксцизионные кольца (TREC) — кольцевые молекулярные структуры, образующиеся в процессе реаранжировки V-генов в результате замыкания в кольцо участков ДНК, вырезаемых из зародышевых V-генов (см. раздел 3.1.4.1). Эти структуры выявляют с помощью полимеразной цепной реакции. Различают сигнальные и кодирующие эксцизионные кольца, образующиеся соответственно при реаранжировке генов 5- и a-цепей (ген TRDV локализован внутри кластера TRAV и может подвергаться перестройке дважды — при построении зрелых генов TRDV и TRAV). Они персистируют в Т-клетках некоторое время после их выхода из тимуса и служат маркером «недавних мигрантов из тимуса». Моноположительные тимоциты обладают функцией соответственно Т-хелперов и цитотоксических Т-лимфоцитов. Однако их функциональная активность выражена слабее, чем у аналогичных клеток периферического отдела иммунной системы (например, при активации они слабее секре- тируют IL-2 и IFNy). Некоторые медуллярные тимоциты, по-видимому, остаются в тимусе и служат источником цитокинов, необходимых для формирования полноценного микроокружения этого органа. Остальные тимоциты покидают тимус через посткапиллярные венулы.
Перемещение созревающих тимоцитов внутри тимуса определяется градиентом хемокинов, секретируемых стромальными (преимущественно эпителиальными) клетками, а также наличием на поверхности тимоцитов соответствующих рецепторов (рис. 3.62). Хотя не все детали этого процесса
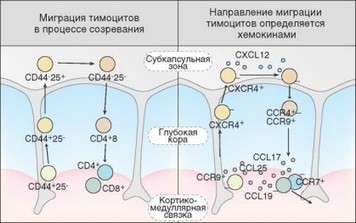
Рис. 3.62. Миграция развивающихся Т-клеток внутри тимуса и зависимость ее направленности от хемокинов. Слева отмечен путь миграции тимоцитов в процессе созревания, справа — хемокины и хемокиновые рецепторы тимоцитов, ответственные за направленный характер перемещений
изучены досконально, известно, что перемещение тимоцитов из кортикомедуллярной в субкапсулярную зоны осуществляется по градиенту SDF-1 (CXCL12), чему способствует возрастающая на этом этапе развития клеток экспрессия соответствующего хемокинового рецептора — CХСR4. На следующем этапе развития тимоциты перемещаются с поверхности коры в глубь ее. Клетки движутся в направлении мест выработки хемокина TECK (ССЫ7), к которому они экспрессируют рецептор CCR9. Экспрессия рецептора CCR7 на заключительном этапе внутритимусного развития Т-клеток обеспечивает их перемещение к кортикомедуллярной зоне. Эндотелиальные клетки венул, через которые осуществляется эмиграция, секретируют хемокин ELC (CCL19), распознаваемый названным CCR7. Рецептор для сфингозина необходим для обеспечения эмиграции тимоцитов в ответ на сигнал от сфингозин-1-фосфата.
Другие клеточные популяции тимуса (см. табл. 3.17)
Помимо тимоцитов в тимусе содержатся другие клетки гематогенного происхождения — макрофаги, В-лимфоциты, дендритные, тучные и миоидные клетки. За исключением дендритных и миоидных, упомянутые клетки не отличаются по свойствам и функции от аналогичных клеток вне тимуса.
Дендритные клетки составляют один из наиболее важных в функциональном отношении типов клеток вилочковой железы. Популяция дендритных клеток тимуса содержит примерно равные количества миелоидных и лимфоидных (плазмоцитоидных) вариантов. Большинство из них дифференцируется в тимусе из предшественников, общих для них и тимоцитов, однако некоторые дендритные клетки тимуса, как полагают, имеют внетимусное происхождение. Основная функция миелоидных дендритных клеток тимуса состоит в осуществлении отрицательной селекции и формировании центральной аутотолерантности. Считают, что плазмоцитоидные дендритные клетки в тимусе, как и вне его, секретируют IFNa, однако роль этого цитокина в тимусе точно не установлена.
Миоидные клетки обладают свойствами, характерными для мышечных и, в меньшей степени, эпителиальных клеток. Они содержат десминовые филаменты, экспрессируют мышечные белки миоглобин, тропонин, рас- пин, синаптофизин . На их поверхности присутствуют рецепторы для ацетилхолина. Миоциты локализуются, главным образом в окружении телец Гассаля и внутри них. Интерес к этой немногочисленной клеточной популяции связан с тем, что они вовлекаются в патологию при тяжелой миастении (миастении гравис), часто сопровождающейся развитием тимом — опухолей, растущих из эпителиальных клеток тимуса.
Автономия тимуса. Возрастная инволюция
Тимус — в значительной степени автономный орган. Он слабо реагирует на потребности периферического отдела иммунной системы (например, на разрушение периферических Т-клеток) и всегда работает с постоянной интенсивностью. При пересадке в один организм нескольких тимусов каждый из них после временной утраты тимоцитов регенерирует до размера, свойственного тимусу соответствующего возраста. Тимус очень чувствителен к действию различных факторов, вызывающих апоптоз клеток. Лимфоидные клетки тимуса погибают под влиянием ионизирующей радиации, введения стероидных гормонов и других повреждающих воздействиях. Его инволюция происходит при стрессе и различных заболеваниях (например, при хронических инфекционных заболеваниях и развитии опухолей). Последствия этой, так называемой акцидентальной (т.е. случайной) инволюции тимуса полностью устраняются регенерацией за счет размножения внутритимусных резервных клеток-предшественников (при этом образуются в основном у5Т-клетки), а затем — за счет размножения и развития клеток-предшественников, мигрирующих из костного мозга.
Важная особенность тимуса — его возрастная инволюция (рис. 3.63). Максимальный относительный вес тимуса выявляют в возрасте 1 года, а максимальный абсолютный вес — в возрасте, предшествующем и сопутствующем половому созреванию (12—14 лет). Затем происходит неуклонное снижение веса тимуса. Ежегодно в молодости теряется около 3%, в более пожилом возрасте — 1% активной ткани тимуса. Инволюция затрагивает в первую очередь корковый слой (с 13—15 лет).
Значительно уменьшается численность тимоцитов и секреторная активность эпителиальных клеток тимуса. Их способность размножаться в клеточной культуре утрачивается еще раньше — вскоре после рождения. В возрасте 23—25 лет инволюция затрагивает мозговой слой тимуса. При этом уменьшается число тимоцитов, эмигрирующих из тимуса в течение суток, и снижается концентрация гормонов тимуса. При сниженной функциональной активности эпителиальных клеток тимуса их жизнеспособность сохраняется долго, но затем происходит постепенная потеря эпителиаль-

Рис. 3.63. Возрастная инволюция тимуса
ных клеток. Утрата лимфоидных и эпителиальных клеток восполняется развитием жировой ткани. У пожилых людей (в возрасте 70 лет и более) тимус заполнен жировой тканью с вкраплением островков со свойственной для нормального тимуса морфологией. Нарушается кортикомедуллярная структура органа, ослабляется функция кортикомедуллярного барьера, что приводит к миграции в кору тимуса нехарактерных для нее клеток, прежде всего В-лимфоцитов, образующих фолликулы. В местах массовой гибели клеток стромы тимуса формируются кисты.
Несмотря на выраженную атрофию, тимус полностью не утрачивает своих функций, что выражается в продолжающейся на очень низком уровне эмиграции клеток, содержащих эксцизионные кольца, и секреции гормонов тимуса. Расчеты, основанные на экстраполяции темпов инволюции, показывают, что полная утрата структур и функций тимуса теоретически должна произойти к возрасту 120 лет. Считают, что возрастная инволюция тимуса служит основой старения иммунной системы в целом.