
Динамичность популяций лимфоцитов проявляется не только в непрерывном перемещении клеток, но и в постоянном их обновлении. Срок жизни лимфоцитов разных субпопуляций существенно варьирует. Самый короткий срок жизни — у естественных киллеров, он составляет 7—10 сут. Продолжительность жизни В-клеток — несколько недель. Срок жизни различных субпопуляций наивных Т-лимфоцитов очень сильно варьирует и составляет 1000—10 000 сут (т.е. примерно 2,5—30 лет). Срок жизни клеток памяти составляет десятки лет и сопоставим с длительностью жизни человека и животных. Замещение отмирающих клеток происходит в основном за счет их образования de novo в процессе лимфопоэза. Однако интенсивность лимфопоэза несколько ниже интенсивности отмирания лимфоцитов. Разница компенсируется за счет спонтанной (фоновой) пролиферации клеток, происходящей очень медленно (доля лимфоцитов, пребывающих в фазе фоновой пролиферации, в каждый конкретный момент не превышает 1%).
Этот баланс уравновешен на уровне лимфоцитарных ниш: численность клеток определенного типа соответствует объему ниш, которые могут быть заселены этими клетками (рис. 3.76). Выше упоминалось о нишах как участках в лимфоидных органах, в которые могут мигрировать В- и Т-лимфоциты, и в которых эти клетки находят необходимые для выживания факторы. Лимфоцитарные ниши можно определить как микроанатомические структуры периферического отдела иммунной системы, способные привлекать мигрирующие лимфоциты и обеспечивающие их факторами выживания, а также необходимыми питательными и энергетическими ресурсами. Внутри популяции происходит конкуренция за места расселения и ресурсы (рис. 3.77).
Если клетки занимают разные ниши, конкуренция отсутствует. В связи с этим важно знать, какие субпопуляции клеток занимают общие и какие —
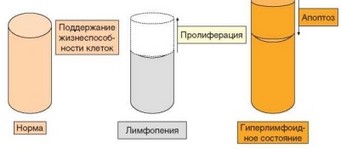
Рис. 3.76. Принцип функционирования гомеостатических лимфоцитарных ниш. Окрашенные цилиндры обозначают ниши, которые могут быть заполнены, недоза- полнены и переполнены. Реакция гомеостатических систем направлена на восстановление соответствия числа клеток размеру ниши
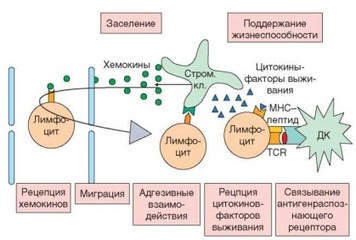
Рис. 3.77. Устройство и принцип функционирования лимфоцитарной ниши. Благодаря наличию хемотаксического сигнала и экспрессии циркулирующими лимфоцитами необходимых молекул адгезии и хемокиновых рецепторов клетки проникают из циркуляции в участок ниши, где они получают необходимые для выживания факторы. Стром. кл. — стромальные клетки
различные ниши. Это устанавливают в опытах с переносом клеток соответствующих субпопуляций в организм, в котором искусственно создается дефицит клеток того или иного типа. Клетки, занимающие общую нишу с находящимися в дефиците клетками, будут в таком случае пролиферировать, чтобы заполнить пустующую нишу. Этот тип пролиферации называют гомеостатической. Ее следует отличать как от фоновой, так и от индуцируемой пролиферации. Так, было установлено, что Т-, В- и NK-клетки зани-
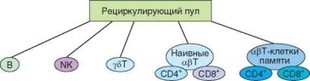
мают разные ниши. Разные ниши занимают у5Т- и арТ-клетки, наивные Т-клетки и Т-клетки памяти (рис. 3.78). Что касается субпопуляций CD4+ и CD8+ Т-лимфоцитов, то они занимают одну нишу, но каждая субпопуляция располагает внутри нее определенной автономией.
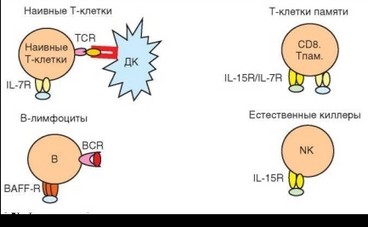
Как правило, факторы, обеспечивающие выживаемость той или иной субпопуляции, одновременно служат индукторами гомеостатической пролиферации, причем неизвестно, почему при нормальном содержании клеток факторы выживания не индуцируют их деления, а при их дефиците активируют пролиферацию клеток недостающего типа, которая прекращается по достижении нормальной их численности. Для всех клеток, кроме наивных Т-лимфоцитов, факторами, контролирующими гомеостаз, служат цитокины (рис. 3.79). Для В-лимфоцитов — это уже упоминашийся выше
BAFF (цитокин семейства TNFa), для естественных киллеров — IL-15 (вспомогательную роль играет IL-7). Гомеостатический эффект IL-15 реализуется через действие не прямо на NK-клетки, а опосредованно через стромальные клетки, связывающие IL-15 своими рецепторами и презенти- рующие его NK-клетке. В поддержании гомеостаза NKT-клеток и Т-клеток памяти также участвуют IL-15 и IL-7.
Гомеостатический контроль наивных Т-клеток реализуется с помощью как гуморальных, так и контактных механизмов. Основную роль в гомеостазе наивных Т-клеток играет IL-7. Варьируя его содержание в организме, можно повышать или понижать численность наивных Т-лимфоцитов. IL-7 вырабатывают стромальные клетки вторичных лимфоидных органов, а также эпителиальные клетки тимуса. При подсадке дополнительных тимусов происходит увеличение численности Т-клеток в результате увеличения содержания IL-7, синтезированного эпителиальными клетками трансплантированного тимуса. При полном отсутствии IL-7 Т-клетки погибают в течение 2—4 нед.
Второй механизм гомеостатического контроля Т-клеток реализуется при помощи того же процесса, что и положительная селекция тимоцитов, только происходит этот процесс не в тимусе, а в периферических лимфоидных органах, прежде всего в паракортикальных зонах лимфатических узлов. Суть его состоит в распознавании TCR арТ-клеток молекул MHC и презентиру- емого ими эндогенного пептида (аналогичного пептидам, распознаваемым при положительной селекции). Для поддержания жизнеспособности и включения гомеостатической пролиферации CD8+ Т-клеток требуется распознавание молекул MHC-I, для гомеостаза CD4+ Т-клеток — молекул MHC-II (рис. 3.80). В последнем случае распознавание может осуществляться только в тимусзависимых зонах лимфоидных органов и только с участием дендритных (интердигитальных) клеток. Для CD8+ Т-клеток условия распознавания MHC-I менее жесткие: считают, что оно может происходить и вне лимфоидных органов, но обязательно с участием дендритных клеток.
Таким образом, зрелые наивные Т-клетки должны постоянно «подтверждать» свою способность «правильно» распознавать антиген. Определенную
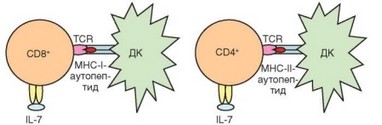
Рис. 3.80. Условия выживания CD8+ и CD4+ Т-клеток. Особое положение наивных Т-лимфоцитов обусловлено зависимостью их выживаемости не только от цитокина (IL-7), но и от прямого распознавания комплексов «молекула MHC—эндогенный пептид» на дендритных клетках
степень автономности гомеостатического контроля этих клеток обеспечивает наличие пределах Т-клеточной ниши гомеостатитеских факторов для CD4+ и CD8+ лимфоцитов в виде разных молекул MHC. При переносе зрелых CD4+ и CD8+ Т-клеток в организм, лишенный молекул MHC одного из классов (например, вследствие генетического дефекта), клетки соответствующей субпопуляции постепенно погибают (CD8+ клетки — через 1—4 нед, CD4+ Т-клет- ки могут прожить до 100 сут). Пока не вполне понятно, как Т-клетки «накапливают» стимул к выживаемости, обусловлено ли это «подсчетом» числа контактов с молекулами MHC и как подобный подсчет осуществляется.
После контакта с антигеном Т-клеткам больше не требуется распознавание молекул MHC для выживания. Жизнеспособность этих клеток, а также контроль их численности осуществляется с участием цитокинов — IL-15, IL-7 и др. Важный аспект гомеостаза лимфоидных популяций — установление равновесия между численностью наивных лимфоцитов и клеток памяти, а также изменение этого баланса с возрастом. Данный вопрос будет рассмотрен далее (см. раздел 3.6.3.2) при описании клеток памяти. Механизмы гомеостатического контроля субпопуляции у5Т-клеток, не способных распознавать молекулы MHC, не установлены. Вероятно, их численность поддерживается только IL-7.
Понимание природы и механизмов гомеостатического контроля численности лимфоцитов в популяциях и субпопуляциях очень важно для правильного истолкования ее изменений при патологиях и разработке путей ее коррекции.