
Кроме первичных иммунодефицитов, единственным заболеванием, для которого поражение иммунной системы является основой патогенеза и определяет симптоматику, является синдром приобретенного иммунодефицита (СПИД; Aquired immune deficiency syndrome — AIDS). Только он может быть признан самостоятельным приобретенным иммунодефицитным заболеванием.
История обнаружения СПИДа восходит к 1981 г., когда в трудах Центра по контролю за заболеваниями (США, штат Атланта) было опубликовано сообщение групп врачей из Нью-Йорка и Лос-Анжелеса о необычном заболевании, зарегистрированном у мужчин-гомосексуалистов. Оно характеризовалось тяжелой формой пневмонии, вызванной условно-патогенным грибком Pneumocystis carinii. В последующих сообщениях были приведены данные о расширении группы больных и приведены данные о наличии у них иммунодефицита, связанного с резким снижением содержания в циркуляции CD4+ Т-лимфоцитов, сопровождающегося развитием инфекционных процессов, которые могут быть вызваны, помимо пневмоцист, другими факультативными патогенами. У некоторых больных развивалась саркома Капоши, характеризовавшаяся несвойственным ей агрессивным течением. К моменту опубликования этих материалов 40% выявленных больных умерли. Позже выяснилось, что эпидемия заболевания уже захватила экваториальную Африку, где болезнь распространяется преимущественно гетеросексуальным половым путем. Международное медицинское сообщество не только признало существование новой нозологической формы — «синдром приобретенного иммунодефицита» (Aquired Immunodeficiency Syndrome), но и констатировало начало пандемии этого заболевания. Столь драматический дебют СПИДа привлек к нему всеобщее внимание, выходящее далеко за пределы профессиональной среды. В медицинской науке, особенно в иммунологии, проблема СПИДа существенно повлияла на распределение усилий и финансов в развитии научных исследований. Это был первый случай, когда заболевание, связанное с преимущественным поражением иммунной системы, оказалось столь значимым в научном и социальном отношениях.
К началу 2007 г. число ВИЧ-инфицированных составило 43 млн, из которых 25 млн погибли, ежегодный прирост этого числа составляет 5 млн, а ежегодная смертность — 3 млн. 60% инфицированных проживают в Африке южнее Сахары.
В 1983 г. почти одновременно во Франции [Л. Монтанье (L. Montagnier)] и Соединенных Штатах Америки [Р.С. Галло (R.C. Gallo)] была определена
вирусная природа СПИД и описан его возбудитель — ВИЧ (вирус иммуно- дифицита человека, Human Immunodeficiency Virus — HIV). Он относится к ретровирусам, т.е. вирусам, у которых носителем наследственной информации служит РНК, и она считывается с участием обратной транскриптазы. Этот вирус принадлежит к подсемейству лентивирусов — медленнодействующих вирусов, вызывающих заболевания с длительным инкубационным периодом. Род ВИЧ включает виды ВИЧ-1, являющийся возбудителем типичной формы СПИДа, и ВИЧ-2, отличающийся от ВИЧ-1 деталями строения и патогенного действия, но в общих чертах аналогичный ему. ВИЧ-2 вызывает более мягкий вариант заболевания, распространенный в основном в Африке. Сведения, приводимые ниже, касаются преимущественно ВИЧ-1 (за исключением особо оговоренных случаев). Выделяют 3 группы ВИЧ — M, O и N, разделяемые на 34 субтипа.
В настоящее время принята точка зрения, что ВИЧ-1 произошел от вируса шимпанзе в Западной Африке (вероятнее всего, в Камеруне — стране, эндемичной по ВИЧ) примерно в 30-е годы ХХ века. ВИЧ-2 произошел от обезьяньего вируса SIVsm. Варианты ВИЧ-1 неравномерно распределены в мире. В развитых странах Запада преобладает субтип В, в центральной Европе и России — субтипы А, В и их рекомбинанты. В Африке и Азии преобладают другие варианты, причем в Камеруне присутствуют все известные субтипы ВИЧ.
Морфология, гены и белки вируса иммунодефицита человека
Схема строения ВИЧ представлена на рис. 4.46. Вирус имеет диаметр около 100 нм. Он окружен оболочкой, из которой выступают грибовидные
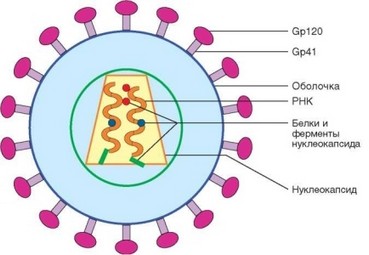
Рис. 4.46. Схема строения вируса иммунодефицита человека 1 (ВИЧ-1)
|
LTR |
gag |
|
vif |
|
vpu |
env |
nef |
LTR |
|||||
|
|
pol |
|
vpr |
|
tat rev |
|
|
||||||
|
|
|
|
? |
|
|
||||||||
Рис. 4.47. Структура генома вируса иммунодефицита человека 1 (ВИЧ-1). Указано расположение генов на двух молекулах РНК вируса шипы, наружная часть которых образована оболочечным белком gp120, а прилегающие к мембране и трансмембранные части — белком gp41. Шипы представляют тримеры названных молекул. Эти белки участвуют во взаимодействии вируса и клетки хозяина, и иммунный ответ последнего направлен в основном против них. Глубже расположен слой матрикса, выполняющий роль каркаса. Срединную часть вируса образует конусообразный капсид, в котором содержится геномная РНК. Здесь же локализованы нуклеопротеиды и ферменты: обратная транскриптаза (р66/р51), интеграза (р31—32), протеаза (р10) и РНКаза (р15).
Генетическая структура ВИЧ и кодируемые его генами белки представлены на рис. 4.47. В двух молекулах односпиральной РНК общей протяженностью 9,2 кбаз локализуются 9 генов, кодирующих 15 белков ВИЧ. Последовательности, кодирующие структуры вируса, ограничены с 5’- и З’-концов длинными концевыми повторами (LTR — Long terminal repeats), выполняющими регуляторные функции. Структурные и регуляторные гены частично перекрываются. Основных структурных гена 3 — gag, pol и env. Ген gag детерминирует образование группоспецифических антигенов сердцевины — нуклеоида и матрикса. Ген pol кодирует ДНК-полимеразу (обратную транскриптазу) и другие белки нуклеотида. Ген env кодирует образование упоминавшихся выше белков оболочки. Во всех случаях первичный продукт генов подвергается процессингу, т.е. расщепляется на более мелкие белки. Регуляторные гены располагаются между генами pol и env (гены vif, vpr, vpu, vpx, rev, tat) и, кроме того, занимают З’-концевую часть генома (фрагменты генов tat и rev, ген nef). Белки, кодируемые регуляторными генами, важны для формирования вириона и его взаимоотношений с клеткой. Из них наиболее важны белки tat — трансактиватор транскрипции и nef (27 кДа) — ее отрицательный регулятор. Дефектный белок nef выявляют у ВИЧ-инфицированных «долгожителей», у которых отсутствует прогрессия заболевания.
Наиболее важны для иммунологии ВИЧ-инфекции, диагностики и разработки подходов к иммунотерапии СПИДа белки оболочки gp120 и gp41. С геном env связана чрезвычайно высокая вариабельность ВИЧ. Ген содержит 5 константных (С) и пять вариабельных (V) участков; в последних последовательность аминокислот варьирует от одного изолята вируса к другому на 30—90%. Особенно значима для иммуногенности вариабельная петля V3. Частота мутаций в гене env составляет 10-4—10-5 событий на геном за цикл, т.е. на 2—3 порядка выше, чем обычная частота мутаций генов. Значительная часть молекулы занята углеводными остатками.
Инфицирование клеток вирусом иммунодефицита человека
Процесс инфицирования клеток человека ВИЧ и его последующей репликации включает несколько стадий. В ранней фазе жизненного цикла можно выделить следующие фазы:
- связывание ВИЧ с поверхностью клетки (рецепция);
- слияние мембран вируса и клетки и проникновение вируса внутрь клетки (фузия и «раздевание»);
- начало обратной транскрипции; формирование преинтеграционного комплекса;
- транспорт преинтеграционного комплекса в нуклеоплазму;
- интеграция провируса в геном клетки.
К этапами поздней фазы жизненного цикла ВИЧ относят:
- транскрипцию вирусной РНК на матрице интегрированной провирусной ДНК;
- экспорт вирусной РНК в цитозоль;
- трансляцию вирусной РНК, процессинг белков;
- сборку вирусной частицы на клеточной мембране;
- высвобождение вновь образованного вириона.
Основные входные ворота инфекции — слизистые оболочки мочеполового и пищеварительного тракта. Проникновение вируса в организм существенно облегчается при наличии повреждений слизистой оболочки, однако инфицирование возможно и при их отсутствии. В этом случае вирус захватывается отростками дендритных клеток, проникающими в просвет органа. В любом случае дендритные клетки первыми взаимодействуют с ВИЧ. Они транспортируют вирус в регионарный лимфоузел, где он в процессе взаимодействия дендритных клеток с Т-лимфоцитами при презентации антигенов инфицирует CD4+ Т-клетки.
Рецепция ВИЧ обусловлена взаимным распознаванием тримера белка gp120 вируса и мембранного гликопротеина CD4 клетки хозяина. На обеих молекулах локализованы участки, ответственные за их взаимодействие. На молекуле gp120 указанный участок расположен в его С-концевой части (остатки 420—469), кроме того, есть еще 3 участка, важные для формирования сайта взаимодействия с CD4, и участок (254—274), ответственный за проникновение вируса в клетку после связывания с мембранным CD4. На молекуле CD4 участок связывания с gp120 расположен в N-концевом V-домене (D1) и включает последовательности остатков 31—57 и 81—94.
Поскольку рецептором для ВИЧ служит молекула CD4, спектр клеток- мишеней этого вируса определяется ее экспрессией (табл. 4.20). Естественно, что главными мишенями для него служат CD4+ Т-лимфоциты, а также незрелые тимоциты, экспрессирующие оба корецептора (CD4 и CD8). Дендритные клетки и макрофаги, слабо экспрессирующие CD4 на мембране, так же эффективно заражаются вирусом и служат его активными продуцентами (репликация ВИЧ в дендритных клетках даже выше, чем в Т-лимфоцитах). Мишенями ВИЧ служат и другие клетки, содержащие на поверхности хотя бы небольшие количества CD4 — эозинофилы, мегакариоциты, эндотелиальные клетки, некоторые эпителиальные (эпителий тимуса, М-клетки кишечника) и нервные клетки (нейроны, клетки микроглии, астроциты, олигодендроци- ты), сперматозоиды, клетки хорионаллантоиса, поперечно-полосатых мышц.
Таблица 4.20. Состояние иммунологических показателей при синдроме приобретенного иммунодефицита
|
Показатель |
Доклиническая стадия |
Стадия клинических проявлений |
|
Число лимфоцитов |
В норме |
Снижено |
|
Содержание CD4+ Т-клеток |
В норме или снижено |
Меньше 200 клеток в 1 мкл крови |
|
Содержание CD8+ Т-клеток |
В норме или повышено |
В норме или снижено (процент может быть повышен) |
|
Соотношение CD4+/CD8+ |
0,8—1,0 |
0,1-0,6 |
|
Соотношение Th1/Th2 |
В норме или снижено |
Снижено |
|
Активность цитоткоси- ческих Т-клеток |
Повышена |
Снижена |
|
Ответ Т-клеток на митогены |
В норме или снижен |
Резко подавлен |
|
Содержание В-клеток |
В норме или снижено |
Снижено |
|
Антигенемия |
Проявляется на 2-8-й неделе |
Осутствует |
|
Антитела в циркуляции |
Появляются обычно после 8 нед |
Присутствуют |
|
Растворимые факторы в циркуляции |
Растворимые формы a-цепи IL-2R, CD8, TNFR, Р2-микроглобулина, неоптерина |
|
|
Содержание и активность NK-клеток |
В норме |
Функция снижена |
|
Лимфоидные ткани, ассоциированные со слизистыми оболочками |
Раннее снижение содержания CD4+ Т-клеток памяти |
Сильное подавление Т-клеток, особенно субпопуляции CD4+ |
|
Врожденный иммунитет |
В норме или подавлен |
Подавлен |
Дополнительные молекулы, необходимые для проникновения ВИЧ в клетки, его корецепторы — 2 хемокиновых рецептора: СХСR4 (рецептор для хемокина CXCL12) и CCR5 (рецептор для хемокинов CCL4 и CCL5). В меньшей степени роль корецептора присуща еще более, чем десятку хемокиновых рецепторов. OXCR4 служит корецептором для штаммов ВИЧ-1, культивируемых на Т-клеточных линиях, а CCR5 — для штаммов, культивируемых на линиях макрофагов (он присутствует на макрофагах, дендритных клетках, а также на CD4+ Т-клетках). Оба эти рецептора относят к родопсиноподобным, передающим в клетку сигнал через связанный с ними G-белок (см. раздел 4.1.1.2). Оба хеморецептора взаимодействуют с белком gp120; участок связывания с этими рецепторами открывается в молекуле gp120 после взаимодействия с CD4 (рис. 4.48). Различные изоля- ты ВИЧ отличаются по избирательности к тем или иным корецепторам. Вспомогательную роль в рецепции ВИЧ-2 играют молекулы адгезии, в частности LFA-1. При инфицировании дендритных клеток во взаимодействии с ВИЧ принимает участие лектиновый рецептор DC-SIGN.
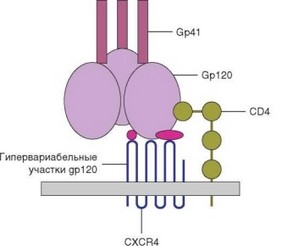
Рис. 4.48. Схема взаимодействия вируса и клетки-мишени при ее инфицировании. Проиллюстрирован один из вариантов взаимодействия рецепторных молекул Т-клетки и молекул ВИЧ-1, обеспечивающего проникновение вируса в клетку
Корецепторам принадлежит важная роль в слиянии вирусной оболочки с мембраной клетки. Со стороны вируса главную роль в слиянии играет белок gp41. После фаз слияния (фузии) и «раздевания» вируса формируется ревертазный комплекс, обеспечивающий обратную траскрипцию с образованием двуспиральной провирусной ДНК.
С помощью вирусного фермента интегразы кДНК интегрируется в ДНК клетки, образуя провирус. Особенность интеграции генов ВИЧ в клеточный геном состоит в том, что для ее осуществления не требуется деление клетки. В результате интеграции формируется латентная инфекция, в которую обычно вовлекаются Т-клетки памяти, «дремлющие» макрофаги, служащие резервом инфекции.
Репликация ВИЧ осуществляется преимущественно или исключительно в активированных клетках. При активации CD4+ Т-клетки происходит индукция транскрипционного фактора NF-KB, который связывается с промоторами как клеточной, так и вирусной ДНК. Клеточная РНК-полиме- раза транскрибирует вирусную РНК. Ранее других транскрибируются гены tat и rev, продукты которых участвуют в репликации вируса. Tat — белок, взаимодействующий c длинными терминальными последовательностями (LTR), что резко повышает скорость вирусной транскрипции. Rev — белок, способствующий выходу из ядра вирусных мРНК-транскриптов, как сплай- сированных, так и не прошедших сплайсинг. Вирусная мРНК, вышедшая из ядра, служит матрицей для синтеза структурных и регуляторных белков. Структурные белки gag, env, pol формируют вирусную частицу, которая отпочковывается от клетки.
Стимуляция лимфоцитов митогенами усиливает репликацию ВИЧ и его цитопатогенный эффект. Этому могут способствовать эндогенные факторы, сопутствующие активации клеток, индуцируемые в активированных лимфоцитах и макрофагах (о NF-kB уже упоминалось). Таким факторами могут быть также цитокины, особенно TNFa и IL-6. Первый активирует транскрипцию генов ВИЧ, второй стимулирует экспрессию ВИЧ в клетках хозяина. Аналогичный эффект оказывают колониестимулирующие факторы GM-CSF и G-CSF. В качестве кофакторов активации ВИЧ могут выступать IL-1, IL-2, IL-3 и IFNy. Глюкокортикоидные гормоны надпочечников способствуют реализации генетической программы ВИЧ. IL-4, IL-7 и IFNa оказывают противоположное действие.
Иммунный ответ на антигены ВИЧ
Острая вирусная инфекция характеризуется сравнительно быстрым образованием антигенспецифичных CD4+ и CD8+ Т-клеток, синтезирующих IFNy. Это приводит к быстрому падению содержания вируса в крови, но не его исчезновению. Клеточный ответ на ВИЧ-инфекцию складывается из образования антигенспецифических CD4+ Т-хелперов и CD8+ Т-кил- леров. Цитотоксические CD8+ Т-клетки выявляют на протяжении всего заболевания СПИДом за исключением поздних стадий, тогда как вирусспецифические CD4+ Т-клетки выявляются только на ранних стадиях заболевания. CD8+ Т-киллеры убивают зараженные клетки до выхода вируса из клетки, прерывая тем самым репликацию вируса. Есть четкая обратная зависимость между титром вируса в плазме крови и количеством специфических CD8+ Т-киллеров. Повышение пролиферативной активности CD4+ и CD8+ антигенспецифических Т-клеток коррелирует с замедлением прогрессирования заболевания. Для больных, содержащих большое количество CD8+ Т-киллеров, характерно медленное прогрессирование заболевания. CD4+ Т-клетки также играют важную роль в элиминации вируса: имеется зависимость между пролиферативным ответом CD4+ Т-клеток на антигены ВИЧ и уровнем вируса в плазме. Отмечено, что выраженность виремии более тесно обратно коррелирует с выработкой IL-2, чем IFNy. При хронической вирусной инфекции в количественном отношении эффекторные Т-клетки сохраняются, но они изменяются функционально. Снижается способность CD4+ Т-клеток синтезировать IL-2; ослабляется образование цитотоксических молекул CD8+ Т-клетками. Снижается пролиферативная активность CD8+ Т-клеток, как полагают, в результате снижения продукции IL-2 CD4+ хелперами. Ослаблению противовирусной защиты способствует дифференцировка CD4+ Т-клеток в хелперы Th2-rarn. Даже для спектра цитокинов, синтезируемых CD8+ цитотоксическими Т-лимфоцитами, характерно преобладание ^2-цитокинов.
Естественно было бы ожидать, что иммунные процессы, которые, пусть и в ослабленной форме, развиваются в ответ на внедряющийся вирус, смогут хотя бы в малой степени защитить организм от инфекции. В действительности, если это и происходит, то лишь на начальном периоде заболевания. В дальнейшем, несмотря на присутствие антигенспецифических CD4+ и CD8+ Т-клеток, происходит интенсивная репликация вируса. Это является следствием отбора вирусов с измененями в эпитопах, распознаваемых
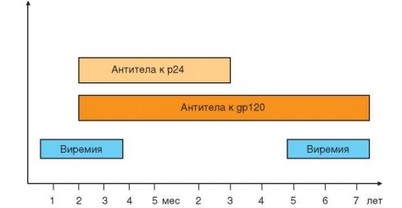
Рис. 4.49. Динамика содержания в крови инфицированных вирусом иммунодефицита человека самого вируса и антител к двум его белкам
Т-клетками, что позволяет им избегать давления со стороны Т-клеточного иммунитета. Таким образом, клеточный иммунный ответ не способен элиминировать вирус из организма в связи с высокой приспособляемостью вируса, основанной на изменчивости. Неэффективны оказываются и NK-клетки, хотя они не являются объектом прямого инфицирования вирусом.
Отражением взаимоотношений между ВИЧ-инфекцией и макроорганизмом служит динамика содержания в циркуляции вирусных антигенов и антивирусных антител (рис. 4.49). Всплеск антигенемии в ранний период развития ВИЧ-инфекции (2—8 нед после инфицирования) отражает интенсивную репликацию вирусов, внедрившихся в клетки. При сохранной иммунной системе хозяина это вызывает наработку нейтрализующих антител (преимущественно к поверхностным белкам gp120, gp41, группоспецифическому gag-антигену р17), что можно выявить по подъему титра сывороточных антител к указанным антигенам, начиная с 8-й недели от момента заражения. Такую смену циркуляции антигена на присутствие в кровотоке антител обозначают термином «сероконверсия». Антитела к оболочечным (env) белкам стабильно сохраняются в течение всего заболевания, тогда как специфичные к gag антитела исчезают на определенных этапах его развития, и вирусные антигены повторно появляются в кровотоке. Одновременно с накоплением в сыворотке крови антител к вирусным антигенам повышается концентрация всех сывороточных иммуноглобулинов, включая IgE.
Циркулирующие антитела способны нейтрализовать свободный вирус и связывать его растворимые белки. При ответе на gp120 это в наибольшей степени относится к антителам, специфичным к иммунодоминантному эпитопу 303—337, локализованному в 3-м гипервариабельном домене (V3) молекулы. Это подтверждается тем фактом, что пассивно введенные антитела могут предохранить от заражения ВИЧ. Нейтрализующие антитела, особенно направленные против gp120, способны блокировать инфициро
вание клеток. Вероятно, это играет определенную роль в первоначальном сдерживании ВИЧ-инфекции и в какой-то степени обусловливают длительный латентный период, характерный для данного заболевания. В то же время эффекторная активность этих антител ограничена и их защитную роль при ВИЧ-инфекции нельзя считать доказанной.
Формирование иммунодефицита при синдроме приобретенного иммунодефицита (см. табл. 4.20)
Основная причина иммунодефицита при СПИДе — гибель CD4+ T-клеток. Очевидная причина гибели инфицированных клеток — цитопатогенное действие вируса. При этом клетки погибают по механизму некроза вследствие нарушения целостности их мембраны. Так, при заражении ВИЧ клеток крови численность CD4+ Т-клеток, начиная с 3-х суток, резко уменьшается одновременно с высвобождением вирионов в среду. В наибольшей степени страдает популяция СD4+ Т-клеток слизистой оболочки кишечника.
Помимо этого механизма гибели инфицированных клеток при СПИДе выявляют высокий уровень апоптоза. Поражение Т-клеточного звена иммунной системы значительно превосходит ожидаемое на основании оценки числа инфицированных клеток. В лимфоидных органах инфицировано не более 10—15% CD4+ Т-клеток, а в крови это количество составляет только 1%, однако апоптозу подвергается значительно больший процент CD4+ Т-лимфоцитов. Помимо инфицированных, апоптотирует значительная часть неинфицированных вирусом клеток, прежде всего CD4+ Т-лимфоцитов, специфичных к антигенам ВИЧ (до 7% этих клеток). Индукторами апоптоза служат белки gp120 и регуляторный белок Vpr, активные в растворимой форме. Белок gp120 понижает уровень антиапоптотического белка Bcl-2 и повышает уровень проапоптотических белков р53, Bax, Bak. Белок Vpr нарушает целостность митохондриальной мембраны, вытесняя Вс1-2. Происходит выход из митохондрии цитохрома с и активация каспазы 9, что приводит к апоптозу CD4+ Т-клеток, в том числе не инфицированных, но ВИЧ-специфичных.
Взаимодействие вирусного белка gp120 с мембранным гликопротеином CD4+ Т-лимфоцитов служит причиной еще одного процесса, происходящего при ВИЧ-инфекции и участвующего в гибели и функциональной инактивации клеток хозяина — формированию синцития. В результате взаимодействия gp120 и CD4 происходит слияние клеток с формированием многоядерной структуры, не способной выполнять нормальные функции и обреченной на гибель.
Среди клеток, инфицируемых ВИЧ, погибают только Т-лимфоциты и мегакариоциты, подвергаясь цитопатогенному действию или вступая в апоптоз. Ни макрофаги, ни эпителиальные или другие клетки, инфицированные вирусом, не теряют жизнеспособности, хотя их функция может нарушаться. Дисфункцию может вызывать не только ВИЧ как таковой, но и его изолированные белки, например, gp120 или продукт гена tat р14. Хотя ВИЧ не способен вызывать злокачественную трансформацию лимфоцитов (в отличие, например, от вируса HTLV-1), белок tat (р14) участвует в индукции саркомы Капоши при ВИЧ-инфекции.
Резкое снижение содержания CD4+ Т-лимфоцитов — самый яркий лабораторный признак ВИЧ-инфекции и ее эволюции в СПИД. Условная граница содержания этих клеток, за которой обычно следуют клинические проявления СПИД, — 200—250 клеток в 1 мкл крови (в относительных цифрах — около 20%). Соотношение CD4/CD8 на пике заболевания снижается до 0,3 и ниже. В этот период проявляется общая лимфопения с уменьшением содержания не только CD4+, но и CD8+ клеток и В-лимфо- цитов. Ответ лимфоцитов на митогены и выраженность кожных реакций на распространенные антигены продолжает снижаться до полной анергии. К разнообразным причинам неспособности эффекторных Т-клеток элиминировать ВИЧ добавляется высокая мутабельность ВИЧ с образованием все новых эпитопов, не распознаваемых цитотоксическими Т-клетками.
Естественно, что среди иммунологических расстройств при СПИДе доминируют нарушения Т-клеточных и Т-зависимых процессов. К факторам, определяющим эти нарушения, относят:
- снижение числа CD4+ Т-хелперов вследствие их гибели;
- ослабление функций CD4+ Т-клеток под влиянием инфицирования и действия растворимых продуктов ВИЧ, особенно gp120;
- нарушение баланса популяции Т-клеток со сдвигом соотношения Th1/Th2 в сторону Th2, тогда как защите от вируса способствуют Thl-зависимые процессы;
- индукция регуляторных Т-клеток белком gp120 и ВИЧ-ассоциирован- ным белком р67.
Снижение способности организма к иммунной защите затрагивает как ее клеточные, так и гуморальные факторы. В результате формируется комбинированный иммунодефицит, делающий организм уязвимым к инфекционным агентам, в том числе условно-патогенным (отсюда — развитие оппортунистических инфекций). Дефицит клеточного иммунитета играет определенную роль в развитии лимфотропных опухолей, а сочетание иммунодефицита и действия некоторых белков ВИЧ — в развитии саркомы Капоши.
Клинические проявления иммунодефицита при инфекции вирусом иммунодефицита человека и синдроме приобретенного иммунодефицита
Основные клинические проявления СПИДа состоят в развитии инфекционных заболеваний, главным образом, оппортунистических. Наиболее характерны для СПИДа следующие заболевания: пневмонии, вызываемые Pneumocystis carinii; диарея, вызываемая криптоспоридиями, токсоплазмами, жиардиями, амебами; стронгилоидоз и токсоплазмоз головного мозга и легких; кандидоз полости рта и пищевода; криптококкоз, диссеминированный или локализованный в ЦНС; кокцидиомикоз, гистоплазмоз, мукормикоз, аспергиллез различной локализации; инфекции нетипичными микобактериями различной локализации; сальмонеллезная бактериемия; цитомегало- вирусная инфекция легких, ЦНС, пищеварительного тракта; герпетическая инфекция кожи и слизистых оболочек; инфекция вирусом Эпштейна-Барр; мультифокальная паповавирусная инфекция с энцефалопатией.
Другую группу связанных со СПИДом патологических процессов составляют опухоли, отличие которых от неассоциированных со СПИДом, состоит в том, что они развиваются в более молодом возрасте, чем обычно (до 60 лет). При СПИДе часто развиваются саркома Капоши и неходжкинс- кие лимфомы, локализующиеся преимущественно в головном мозгу.
Развитию патологического процесса способствуют некоторые реакции макрооргнаизма, провоцируемые ВИЧ-инфекцией. Так, активация CD4+ Т-клеток в ответ на действие вирусных антигенов способствует реализации цитопатогенного эффекта, особенно апоптоза Т-лимфоцитов. Большинство образуемых при этом Т-клетками и макрофагами цитокинов благоприятствуют прогрессированию ВИЧ-инфекции. Наконец, в патогенезе СПИДа важную роль играет аутоиммунная составляющая. Ее основу составляет гомология между белками ВИЧ и некоторыми белками организма, например между gp120 и молекулами MHC. Однако эти нарушения, усугубляя иммунодефицит, не формируют специфических аутоиммунных синдромов.
Уже на доклинической стадии ВИЧ-инфекции возникает необходимость использования иммунологичеких методов диагностики. С этой целью используют иммуноферментные тест-наборы, позволяющие определять наличие в сыворотке крови антител к белкам ВИЧ. Существующие тестсистемы основаны на твердофазном иммуносорбентном тестировании антител (ELISA). Первоначально применяли тест-наборы с использованием в качестве антигенного материала вирусных лизатов. Позже с этой целью стали применять рекомбинантные белки ВИЧ и синтетические пептиды, воспроизводящие эпитопы, с которыми взаимодействуют сывороточные антитела ВИЧ-инфицированных людей.
В связи с исключительно высокой ответственностью врачей, делаю - щих заключение об инфицированности ВИЧ на основании лабораторных анализов, принята практика повторной постановки анализов на антитела (иногда с помощью альтерантивных методов, например иммуноблоттинга, см. раздел 3.2.1.4), а также определение вируса с помощью полимеразной цепной реакции.
Лечение СПИДа основано на применении противовирусных препаратов, среди которых наиболее широко используют зидовудин, действующий как антиметаболит. Успехи достигнуты в контроле течения СПИДа, существенно увеличивающем продолжительность жизни больных. Основной терапевтический подход — использование антиметаболитов нуклеиновых кислот в варианте высокоактивной антиретровирусной терапии (High active antiretroviral therapy — HAART). Эффективным дополнением к антиретровирусной терапии служит применение препаратов интерферонов, а также лечение сопутствующих заболеваний и вирусных инфекций, способствующих прогрессированию СПИД.
Летальность от СПИД до сих пор составляет 100%. Наиболее частой причиной смерти являются оппортунистические инфекции, особенно пневмоцистные пневмонии. Другие причины смерти — сопутствующие опухоли, поражение центральной нервной системы и пищеварительного тракта.