
 Эритроциты - самая многочисленная популяция клеток крови. Эритроцит в покое имеет форму двояковогнутого диска, которая увеличивает его площадь и обеспечивает наибольшую поверхность газообмена (рис. 5).
Эритроциты - самая многочисленная популяция клеток крови. Эритроцит в покое имеет форму двояковогнутого диска, которая увеличивает его площадь и обеспечивает наибольшую поверхность газообмена (рис. 5).
Диаметр наибольшего числа эритроцитов составляет
- 7,5 мкм, площадь поверхности - 140 мкм2. объем - 90 мкм3 (фл). Высокая пластичность и деформируемость мембраны позволяют эритроциту проходить через капилляры диаметром 2-5 мкм, через стенки синусоидов, диаметр пор которых в селезенке имеет 0,5-0,7 мкм, и затем возвращаться к исходным параметрам.
Клетка покрыта двухслойной полупроницаемой мембраной (плазмолем- мой), в двойной слой фосфолипидов которой встроены белки гликопротеины, углеводная часть их образует надмембраиный слой - гликокаликс. Клетка имеет спектрин-актиновый цитоскелет, большая часть белков которого также составляют фосфолипиды. Клеточная мембрана и цитоскелет определяют архитектонику эритроцита и его способность к деформации при прохождении микроциркуляторного русла. Гликопротеиновые комплексы мембраны организованы таким образом, что отрицательно заряженные участки (чаще сиаловые группы полисахаридов) обращены наружу, придавая поверхности эритроцитов отрицательный заряд.
Внутренняя сторона мембраны эритроцитов связана с сетью миофиламент- ных белков, формирующих спектрин-актиновый цитоскелет, придающий эритроциту специфическую двояковогнутую форму (рис. 6).
В ячейках цитоскелета находятся молекулы гемоглобина. По данным
Т.С. Истомановой и соавт., число молекул НЬ в 1 эритроците составляет около 280 млн, по данным А.А. Липаца - 400 млн.
Гемоглобин (НЬ) - дыхательный пигмент, сложный белок - хромопротеид. Его небелковая часть (простетическая группа), включающая железо, называется гемом, белковый компонент - глобином. На долю глобина приходится 96% сухого веса НЬ, на долю гема - 4%. Молекула НЬ имеет 4 гема. Благодаря присутствию в составе гема иона железа, гемоглобин переносит кислород от легочных альвеол к тканям и осуществляет транспортировку углекислого газа от тканей к легким.
Синтез гемоглобина начинается на самой ранней стадии развития эритроидных элементов. При его нарушении содержание НЬ в эритроците
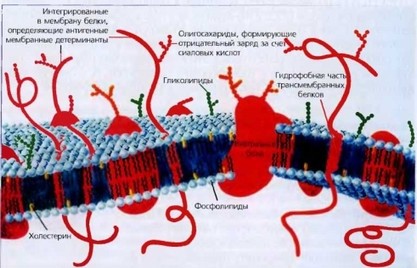
Рис. 6. Схема мембраны эритроцита
снижается, ячейки цитоскелета остаются не заполнены НЬ, что проявляется гипохромией эритроцитов в мазках крови и повышением в них концентрации неиспользованного на синтез порфирина. Мм гемоглобина - 64-66 кДа. В эритроцитах взрослых людей 95-98% приходится на Hb A (adult - взрослый), 2-3%-наНЬ А2,1-2%-HaHbF (fetus-плод). Гемоглобин? у новорожденных составляет 70-90%, но к концу первого года жизни его количество резко снижается. HbF может присутствовать не во всех эритроцитах.
Эритроцитарные антигены. На мембране эритроцита имеется более 250 антигенов, которые располагаются подобно мозаике. Роль антигенов заключается в регуляции дифференцировки и созревания клеток. Наиболее изучены антигены систем АВО и резус. Резус-антигены находятся на мембране эритроцитов независимо от пола и возраста, не связаны с изоантигенами А, В и др., равномерно распределяясь во всех группах крови. Эритроцитарные антигены не меняются в течение жизни человека и могут тысячелетиями сохраняться у трупов при низких температурах и неопределенно долго при высыхании. Распад клетки приводит к потере ею специфической антигенности. Системы эритроцитарных антигенов наследуются обычно независимо друг от друга. Ген, определяющий группу крови системы АВО, обладает тремя аллелями - в виде антигенов А, В и отсутствия этих антигенов в виде третьей формы - 0. Антигены А и В являются кодоминантными и доминируют над 0-антигеном. Резус-антигены при передаче их от обоих родителей могут проявляться в двойной дозе (гомозиготы) или в простой дозе (гетерозиготы) при передаче от одного из родителей. Например, антигены группы А (II) - в виде АА или АО, резус-антигены - CDE/CDE или CDE/cDe. Антигены А и В впервые обнаруживаются на 37-й день развития эмбриона. Формирование резус-антигенов начинается с 3—4-го месяца внутриутробного развития.
Мембрана эритроцита выполняет разные функции: барьерную, транспортную, сорбционную (особенно в отношении микроэлементов), метаболическую благодаря наличию ферментных систем, регулирующих энергетические и окислительные процессы, транспорт ионов, перекисное окисление липидов, генерирующих и утилизирующих активные формы кислорода.
Метаболизм эритроцита. Эритроцит переносит кислород, но сам его не использует для образования собственной энергии, потому что к моменту созревания клетки он теряет митохондрии и рибосомы, в нем исчезают процессы, связанные с образованием энергии путем утилизации кислорода. АТФ в эритроцитах образуется исключительно в процессе анаэробного гликолиза, при этом очень активно используется глюкоза. Эритроциты, составляя около 3% массы тела, в день потребляют приблизительно 20 г (10%) всего количества глюкозы, поступающей в организм. Процесс гликолиза ферментативный, в нем участвует более десяти ферментов. При их недостаточности эритроциты быстро разрушаются. В процессе гликолиза формируются продукты (соединения), которые снижают сродство гемоглобина к кислороду, способствуя отдаче его тканям. Следующие метаболические пути обеспечивают функциональную активность эритроцитов: 1) гликолиз; 2) пентозофосфатный цикл; 3) путь Раппопорта-Либеринга - образования 2,3-дифосфоглицерата (2,3-ДФГ); 4) глутатионовый путь; 5) АТФ-зависимый катионный насос на мембране эритроцита; 7) ферменты, влияющие на состав фосфолипидов эритроцитарной мембраны; 8) метгемоглобинредуктаза, превращающая метгемог- лобин (MetHb) в гемоглобин (НЬ).
Последовательность основных этапов гликолиза и ферменты, катализирующие этот процесс, представлены на рис. 7.
В эритроцитах, в отличие от других тканей, в процессе гликолиза образуется много 2,3-дифосфоглицериновой кислоты, которая, соединяясь с В-цепями глобина, снижает сродство гемоглобина к кислороду, способствуя тем отдаче его тканям.
Из всех ферментов, обеспечивающих гликолиз, чаще других встречается наследственный дефицит пируваткиназы, которая катализирует его энергообразующую стадию, реже глюкозофосфатизомеразы. Нарушения активности других ферментов очень редки. Конечные продукты гликолиза - молочная
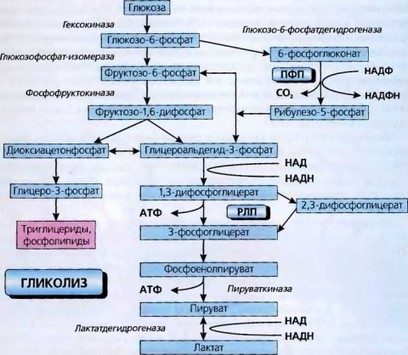
Рис. 7. Гликолиз и сопряженные с ним метаболические процессы, протекающие в эритроцитах: ПФП — пентозофосфатный путь (шунт); РЛП — Раппопорта-Либеринга путь образования 2,3-ДФГ
кислота и АТФ. Прекращение гликолиза приводит эритроцит к «метаболической смерти» - процессу, который заканчивается гемолизом.
Кроме того, до 30% глюкозы эритроцита используется в пентозофос- фатном цикле, результатом которого является синтез НАДФН2 - основного метаболита, обеспечивающего функционирование антиоксидантных систем в эритроците. Эта система эритроцита предупреждает чрезмерное накопление свободных радикалов двухвалентного железа и кислорода, способных вызвать активацию перекисного окисления липидов эритроцитарной мембраны и его гемолиз.
Избыточное образование продуктов перекисного окисления и глики- рование клеточных структур оказывают цитотоксическое повреждающее действие на мембраны эритроцитов и эндотелий сосудов. Конечные продукты гликирования необратимо связываются с белками клеток, изменяя их биологические и функциональные свойства, снижая способность эритроцитов к деформации, что в свою очередь приводит к застойным явлениям в микроциркуляторном русле и, как следствие, к возникновению тканевой гипоксии с выраженными изменениями гомеостаза и гемодинамики. Глико- зилирование белковых структур сосудистой стенки усиливает образование эндотелина-1, обладающего вазоконстрикторным (сосудосуживающим) действием. Дизрегуляция ригидности сосудистой стенки и увеличение внутрисосудистого давления обусловлены также дисфункцией мембран эритроцитов, нарушением их способности сорбировать и транспортировать в ткани норадреналин и ацетилхолин, участвующие в регуляции тонуса сосудистой стенки.
Нарушение доставки кислорода крови тканям рассматривается как показатель тяжести патологического процесса и метаболических изменений в эритроцитах и в организме, поскольку эритроциты ответственны за кислородное обеспечение всех энергообразуюгцих в нем процессов.