
Объективная регистрация состояния бодрствования с помощью электроэнцефалографии стала в наше время одной из повседневных процедур нейробиологического исследования и клинической неврологии (рис. 39) (см. стр. 24).
Альфа-волны. Основным типом волн у нормальных взрослых людей являются альфа-волны, представляющие собой кривую, близкую к синусоиде. Впервые такую активность описал Бергер (Berger). Частота альфа-ритма составляет 8—13 Гц. Лучше всего он отводится от затылочной области и характерен
4
5
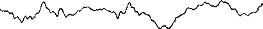
Рис. 39. Основные волны ЭЭГ: 1 — бета-волны; 2 — альфа-волны; 3 — альфа- и тэта-волны; 4 — тэта- и дельта-волны; 5 — дельта-волны.
для людей, находящихся в состоянии физического и психического расслабления, с закрытыми глазами.
Бета-волны. Для активной сенсорной деятельности характерны бета-волны частотой 13—30 Гц, с амплитудой ниже, чем у альфа-волн, т. е. в диапазоне от 5 до 50 мкВ. Бета-волны вызываются разными факторами (например, открыванием глаз, фиксацией взора, звуковыми или кожными стимулами или иными активирующими воздействиями). Бета-ритм регистрируется в лобных областях в состоянии покоя.
Дельта-волны. Основной формой ритмической активности во время сна являются дельта-волны. Их частота равна 4 Гц, а амплитуда обычно превышает 50 мкВ. Они регистрируются у нормальных детей в возрасте до 10 лет во время бодрствования и у взрослых во время сна.
Тэта-волны. У этих волн, промежуточных между альфа- и дельта-волнами, амплитуда выше, чем у первых, и ниже, чем
у вторых. В норме они регистрируются у взрослых в дремоте перед пробуждением или при засыпании.
Всем этим типам волн свойственна большая индивидуальная изменчивость даже при одинаковых условиях отведения. Однако здесь все же возможны некоторые обобщения.
В раннем возрасте доминируют высокоамплитудный дельтаритм и высокочастотный бета-ритм. ЭЭГ взрослого типа формируется после полового созревания. В затылочных отведениях частота всегда ниже, а амплитуда выше, чем в лобных. Электрическая активность в обоих полушариях приблизительно одинакова, если она регистрируется в симметричных точках. Частота снижается (высокая амплитуда, низкая частота) при снижении уровня кислорода или повышении уровня С02 в крови и возрастает, когда содержание кислорода в крови увеличивается, а содержание С02 падает.
Наиболее явная корреляция обнаружена между формой ЭЭГ и тонусом больших полушарий. Амплитуда находится в обратной, а частота — в прямой зависимости от уровня бодрствования. Быстрая активность характерна для бодрствующего мозга. Переход от альфа- к бета-активности наблюдается при яркой вспышке света или в том случае, если человеку предлагают сосредоточиться на решении умственной задачи (например, произвести сложение). Это явление называется блокадой альфа-ритма или десинхронизацией. Чем большая часть мозга заторможена, т. е. находится в состоянии сна, тем выше амплитуда и тем ниже частота волн. «Медленная активность» характерна для мозга, на который не воздействуют внешние раздражения. Поскольку форма ЭЭГ достаточно верно отражает состояние бодрствования или сна, ее используют при контроле наркоза во время операций.
Применение в клинике. Если форма ЭЭГ противоречит функциональному состоянию человека (например, если у бодрствующих людей появляются дельта-волны), то это надо считать признаком патологии. Применение ЭЭГ в клинике полезно при диагностике некоторых опухолей, абсцессов, эпилептоген- ных очагов и др.
Происхождение волн ЭЭГ. Потенциал, регистрируемый элекгроэнцефалографом, несомненно, является суммой электрической активности большого числа корковых нейронов. Но какого рода потенциалы суммируются? И какой механизм обусловливает регулярность альфа-ритма? На первый вопрос смогут ответить микроэлектродные исследования. Вероятно, каким-то образом суммируются спайки корковых нейронов, и в этом участвуют также локальные постсинаптические потенциалы.
Что касается механизма, организующего потенциалы в характерную форму регулярных волн, то тут возможны только гипотезы. Регулярность «медленной активности» (альфа- и
дельта-ритмов) показывает, что группы нейронов в той области, от которой она отводится, по какой-то причине деполяризуются синхронно.
Синхронизация. Синхронизация, вероятно, контролируется таламусом. При стимуляции ретикулярных клеток таламуса серией электрических импульсов частотой 5—-15 Гц в коре регистрируются волны, сходные с синхронизованными волнами ЭЭГ. Как полагают, эти «волны вовлечения» создаются таламическими ядрами. При стимуляции неспецифических ядер таламуса синхронизованные «волны вовлечения» появляются во всей коре. Поэтому можно думать, что синхронизованные волны в неактивной коре контролируются таламической ретикулярной формацией.
Десинхронизация. Лучше известен механизм «быстрой активности» корковых нейронов. Она не содержит синхронизованных составляющих — наоборот, в это время происходит десинхрони- зацня. Когда ЭЭГ начали использовать в клинике, вскоре было замечено, что внешние стимулы вызывают прекращение альфа-ритма, который переходит в низкоамплитудную бета-ак- тивность. Тот факт, что у экспериментальных животных десинхронизацию можно вызвать электрическим раздражением ретикулярной формации ствола мозга, помог выяснить, каким образом альфа-ритм блокируется сенсорными импульсами. Для бодрствования коры, т. е. для появления бета-волн, необходима интактная ретикулярная формация. Импульсы, поступающие по специфическим афферентным путям, не могут пробудить мозг без участия ретикулярной формации. Все эти данные показывают, что изменение ЭЭГ при пробуждении, т. е. замена медленной синхронизованной дельта-активности более быстрыми альфа-волнами, а затем бета-волнами, происходит благодаря тому, что в процесс постепенно вовлекается ретикулярная формация.
Две сенсорные системы. Как показывают также и волны ЭЭГ, рецепторы сенсорной системы связаны с корой афферентными волокнами двух видов.
Один вид неспецифический, и он участвует в реакции пробуждения, проявляющейся в ЭЭГ. От специфических сенсорных путей идут коллатерали к ретикулярным нейронам; эти волокна оканчиваются в коре диффузно после нескольких переключений. Таким образом стимуляция сенсорных путей вызывает ретикулярную активацию и ускорение ЭЭГ-активности.
Вторая сенсорная система — та, что описана в предыдущих главах; это система собственно сенсорных путей, состоящих из трех пли более нейронов каждый, с четко оформленными таламическими ядрами, высоким уровнем организации и разграниченными корковыми представительствами. Как уже говорилось при описании методов исследования сенсорной деятельно
сти, специфические афферентные сигналы проявляются в соответствующих корковых областях в виде вызванных потенциалов. Таким образом, двум афферентным системам соответствуют два типа электрических реакций, регистрируемых в коре: специфические сенсорные импульсы создают вызванные потенциалы в соответствующих зонах коры, а затем во всей норе возникают бета-волны, означающие ретикулярную активацию.
Альфа-волны. Основным типом волн у нормальных взрослых людей являются альфа-волны, представляющие собой кривую, близкую к синусоиде. Впервые такую активность описал Бергер (Berger). Частота альфа-ритма составляет 8—13 Гц. Лучше всего он отводится от затылочной области и характерен
4
5
Рис. 39. Основные волны ЭЭГ: 1 — бета-волны; 2 — альфа-волны; 3 — альфа- и тэта-волны; 4 — тэта- и дельта-волны; 5 — дельта-волны.
для людей, находящихся в состоянии физического и психического расслабления, с закрытыми глазами.
Бета-волны. Для активной сенсорной деятельности характерны бета-волны частотой 13—30 Гц, с амплитудой ниже, чем у альфа-волн, т. е. в диапазоне от 5 до 50 мкВ. Бета-волны вызываются разными факторами (например, открыванием глаз, фиксацией взора, звуковыми или кожными стимулами или иными активирующими воздействиями). Бета-ритм регистрируется в лобных областях в состоянии покоя.
Дельта-волны. Основной формой ритмической активности во время сна являются дельта-волны. Их частота равна 4 Гц, а амплитуда обычно превышает 50 мкВ. Они регистрируются у нормальных детей в возрасте до 10 лет во время бодрствования и у взрослых во время сна.
Тэта-волны. У этих волн, промежуточных между альфа- и дельта-волнами, амплитуда выше, чем у первых, и ниже, чем
у вторых. В норме они регистрируются у взрослых в дремоте перед пробуждением или при засыпании.
Всем этим типам волн свойственна большая индивидуальная изменчивость даже при одинаковых условиях отведения. Однако здесь все же возможны некоторые обобщения.
В раннем возрасте доминируют высокоамплитудный дельтаритм и высокочастотный бета-ритм. ЭЭГ взрослого типа формируется после полового созревания. В затылочных отведениях частота всегда ниже, а амплитуда выше, чем в лобных. Электрическая активность в обоих полушариях приблизительно одинакова, если она регистрируется в симметричных точках. Частота снижается (высокая амплитуда, низкая частота) при снижении уровня кислорода или повышении уровня С02 в крови и возрастает, когда содержание кислорода в крови увеличивается, а содержание С02 падает.
Наиболее явная корреляция обнаружена между формой ЭЭГ и тонусом больших полушарий. Амплитуда находится в обратной, а частота — в прямой зависимости от уровня бодрствования. Быстрая активность характерна для бодрствующего мозга. Переход от альфа- к бета-активности наблюдается при яркой вспышке света или в том случае, если человеку предлагают сосредоточиться на решении умственной задачи (например, произвести сложение). Это явление называется блокадой альфа-ритма или десинхронизацией. Чем большая часть мозга заторможена, т. е. находится в состоянии сна, тем выше амплитуда и тем ниже частота волн. «Медленная активность» характерна для мозга, на который не воздействуют внешние раздражения. Поскольку форма ЭЭГ достаточно верно отражает состояние бодрствования или сна, ее используют при контроле наркоза во время операций.
Применение в клинике. Если форма ЭЭГ противоречит функциональному состоянию человека (например, если у бодрствующих людей появляются дельта-волны), то это надо считать признаком патологии. Применение ЭЭГ в клинике полезно при диагностике некоторых опухолей, абсцессов, эпилептоген- ных очагов и др.
Происхождение волн ЭЭГ. Потенциал, регистрируемый элекгроэнцефалографом, несомненно, является суммой электрической активности большого числа корковых нейронов. Но какого рода потенциалы суммируются? И какой механизм обусловливает регулярность альфа-ритма? На первый вопрос смогут ответить микроэлектродные исследования. Вероятно, каким-то образом суммируются спайки корковых нейронов, и в этом участвуют также локальные постсинаптические потенциалы.
Что касается механизма, организующего потенциалы в характерную форму регулярных волн, то тут возможны только гипотезы. Регулярность «медленной активности» (альфа- и
дельта-ритмов) показывает, что группы нейронов в той области, от которой она отводится, по какой-то причине деполяризуются синхронно.
Синхронизация. Синхронизация, вероятно, контролируется таламусом. При стимуляции ретикулярных клеток таламуса серией электрических импульсов частотой 5—-15 Гц в коре регистрируются волны, сходные с синхронизованными волнами ЭЭГ. Как полагают, эти «волны вовлечения» создаются таламическими ядрами. При стимуляции неспецифических ядер таламуса синхронизованные «волны вовлечения» появляются во всей коре. Поэтому можно думать, что синхронизованные волны в неактивной коре контролируются таламической ретикулярной формацией.
Десинхронизация. Лучше известен механизм «быстрой активности» корковых нейронов. Она не содержит синхронизованных составляющих — наоборот, в это время происходит десинхрони- зацня. Когда ЭЭГ начали использовать в клинике, вскоре было замечено, что внешние стимулы вызывают прекращение альфа-ритма, который переходит в низкоамплитудную бета-ак- тивность. Тот факт, что у экспериментальных животных десинхронизацию можно вызвать электрическим раздражением ретикулярной формации ствола мозга, помог выяснить, каким образом альфа-ритм блокируется сенсорными импульсами. Для бодрствования коры, т. е. для появления бета-волн, необходима интактная ретикулярная формация. Импульсы, поступающие по специфическим афферентным путям, не могут пробудить мозг без участия ретикулярной формации. Все эти данные показывают, что изменение ЭЭГ при пробуждении, т. е. замена медленной синхронизованной дельта-активности более быстрыми альфа-волнами, а затем бета-волнами, происходит благодаря тому, что в процесс постепенно вовлекается ретикулярная формация.
Две сенсорные системы. Как показывают также и волны ЭЭГ, рецепторы сенсорной системы связаны с корой афферентными волокнами двух видов.
Один вид неспецифический, и он участвует в реакции пробуждения, проявляющейся в ЭЭГ. От специфических сенсорных путей идут коллатерали к ретикулярным нейронам; эти волокна оканчиваются в коре диффузно после нескольких переключений. Таким образом стимуляция сенсорных путей вызывает ретикулярную активацию и ускорение ЭЭГ-активности.
Вторая сенсорная система — та, что описана в предыдущих главах; это система собственно сенсорных путей, состоящих из трех пли более нейронов каждый, с четко оформленными таламическими ядрами, высоким уровнем организации и разграниченными корковыми представительствами. Как уже говорилось при описании методов исследования сенсорной деятельно
сти, специфические афферентные сигналы проявляются в соответствующих корковых областях в виде вызванных потенциалов. Таким образом, двум афферентным системам соответствуют два типа электрических реакций, регистрируемых в коре: специфические сенсорные импульсы создают вызванные потенциалы в соответствующих зонах коры, а затем во всей норе возникают бета-волны, означающие ретикулярную активацию.